






Plant Ecology of the Sonoran Desert Region
You could easily recognize a desert even if you were blind- folded. You would discover that you could walk fairly long distances without bumping into plants, and when you did the encounter would likely be painful. Even standing still you would have unmistakable clues about your location. You�d feel the arid atmosphere pulling moisture out of your body and experience a sensation of pressure on your skin from the intense sunlight. On really hot, dry days you could smell pungent, aromatic terpenes and oils exuded by the parched vegetation.
With the blindfold removed, you would see that most desert plants also look different from those in other habitats�they are often spiny, almost always tiny-leafed, and rarely �leaf green.� Many have bold, sculptural growth forms characterized by swollen stems or starkly exposed stems unconcealed by foliage. At the other extreme is a unique desert growth form that landscape architect Iain Robertson calls �diaphanous plants.� The stems and foliage of these plants are so fine-textured and sparse that the eye tends to look right through them.
These tactile, olfactory, and visual experiences offer clues to desert plants� adaptations to their rigorous environment. Before exploring these special characteristics, it is important that you understand something about plant structures, functions, and classification. ![]()
Basic Plant Anatomy and Classification
Many people mistakenly identify ocotillos, agaves, African euphorbias, and numerous other plants as cacti because of their succulent or spiny stems, when in fact these plants are not related to each other or to cacti. Frequently plants (and animals) are similar to each other in outward appearance because their ancestors have adapted to the same environmental challenges by evolving similar forms or structures. This similarity in response to environment, despite lack of common recent ancestors, is known as convergent evolution. The very similar outward appearance of some New World cacti and Old World succulent euphorbias is an excellent example.
Unlike overall form or vegetative structures, the sexual parts of plants (flowers and fruits) are reliable indicators of interrelationships and means of identification. Flowers must function successfully if a plant is to reproduce. Therefore the floral structures tend to remain more consistent within a species than do vegetative parts. Floral structures also form complex patterns that are more readily traceable as plants evolve. The parts of flowers and fruits are also easier to identify and describe than the vegetative organs (leaves, stems, and roots). Moreover, qualitative vegetative characters are hard to describe precisely even when the overall appearance (gestalt) is distinctive. For example, nearly every hiker knows poison ivy on sight. But try to describe the foliage so precisely that someone who has never seen the plant can distinguish it from skunk bush (Rhus trilobata). It�s quite difficult to delineate the leaves� different shades of green, degrees of hairiness, and the scalloping of the margins, especially if you lack the minutely-detailed vocabulary of the botanist. For example, the terms pubescent, puberulent, lanate, villous, hirsute, hirsutulous, ciliate, tomentose, strigose, pilose, and hispid are just some of those used to describe different kinds and degrees of hairiness. Vegetative parts are also more plastic; that is, they vary greatly�even in the same individual�under environmental influences. The leaves of brittlebush grow much larger and greener in shade or during rainy periods than in sun or in drier conditions. (See photo on page 131; see also species account.)
The complex parts of flowers and fruits are arranged in distinctive patterns that can be characterized exactly. Petals, stamens, and other structures can be counted and their lengths and widths measured (and these are usually less variable than the dimensions of leaves). The point of attachment of the stamens to the petals (or other parts) can be described unambiguously. For example, a flower that has many (more than ten) petals and sepals that intergrade into one another, many stamens (usually hundreds), a two- to multi-lobed stigma, and an ovary enveloped in stem tissue may be unequivocally identified as belonging to a member of the cactus family. All 2000 species of cacti possess some variation of this basic pattern, and no other plant group does.
To recognize floral patterns you must be able to identify the parts of a flower. The drawing on page 131 shows the anatomy of a generalized flower. ![]()
Flower Anatomy, from Outside to Inside
In the game �Twenty Questions� players attempt to identify an unknown by asking the person who knows the answer a series of yes-or-no questions. If done well, twenty questions is sufficient to eliminate every other possibility in the world and leave the correct answer standing. Assume, for example, that the unknown thing is a dog. First question: �Is it a concept?� (No; therefore it�s an object.) Second question: �Is it alive?� (Yes.) Third question: �Is it a plant?� (No). Fourth question: �Is it a vertebrate? (Yes). Fifth question: �Is it an herbivore?� (No.) The enormous inventory of the universe has been narrowed to a very short list in only five questions.
Botanists identify plants (and zoologists, animals) unknown to them with a Twenty Questions-like procedure called a dichotomous key (or simply, key). A key is a nested series of dual choices that quickly narrows the possibilities to a single species. For example, the first pair of choices might ask you whether the flower has three petals versus four or five. Each of the two possible answers leads to another pair of choices, and so on, until you have identified your quarry out of 300,000 species of flowering plants. But before you can use such a key effectively, or before you can describe your unknown to someone who will identify it for you, you must know the parts of the flower and plant you are examining.
The sepals collectively make up the calyx. They enclose all other flower parts in the bud, usually completely concealing the rest of the flower until it opens. The petals collectively make up the corolla. Petals are frequently the visual advertising banner that attracts pollinators. Petals and sepals look similar in many flowers, such as in lilies and agaves. By definition the sepals are the parts on the outside; petals are typically concealed in the bud stage. When sepals and petals can�t be readily distinguished, they are called tepals.
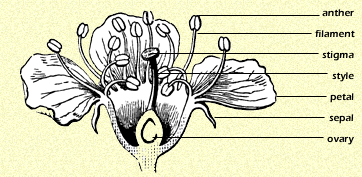 The corolla and calyx make up the perianth. The perianth parts may be separate or fused together for part or all of their length. Often there is only one series of perianth parts. Of necessity these must be on the outside and therefore they are sepals, even if they are large and colorful.
The corolla and calyx make up the perianth. The perianth parts may be separate or fused together for part or all of their length. Often there is only one series of perianth parts. Of necessity these must be on the outside and therefore they are sepals, even if they are large and colorful.
The female part of a flower is the pistil, composed of stigma, style and ovary (also called ovulary). The ovary contains ovules, which develop into seeds when fertilized by the sperm in pollen. Seeds are plant embryos encased in a protective membrane, usually along with stored energy to fuel germination. If the ovary is visible beneath the calyx, it is said to be inferior. It is superior if you must look inside the flower to see it (that is to say, it is above the calyx).
The male part of a flower is the stamen, composed of the anther and the filament. Anthers produce pollen grains, which contain sperm cells. ![]()
Photosynthesis
Chlorophyll is one of the most consequential chemicals in the biosphere. Nearly all life on the planet depends on it. Living organisms seem to defy the law of entropy, the universal tendency toward increasing disorder in a closed system. By using energy acquired from outside they prevent themselves�temporarily�from dying and disintegrating into simple, dissociated molecules (becoming disordered). A small number of species derive their energy from metabolizing sulfur compounds. All others, including all the organisms that we encounter in everyday life, depend on solar energy (light) to maintain their orderly existence. Light, however, is unmanageable; it can�t be concentrated and stored for later use. (Outside of science fiction there is no such thing as a photon battery). Enter photosynthesis. Green plants use light energy to combine low-energy molecules (carbon dioxide and water) into high-energy molecules (carbohydrates), which they accumulate and store as energy reserves. Chlorophyll (the green pigment in plants) is the only known substance in the universe that can capture volatile light energy and convert it into a stable form usable for biological processes (chemical energy).
Almost without exception living organisms�plants, animals, and even fungi and bacteria�obtain energy for sustaining life from carbohydrates (sugars and starches) by the meta- bolic process of aerobic respiration. (�Respiration� is colloquially and medically used to mean breathing. The mechanical act of breathing, however, is only the first step in the physiological process of respiration.) Respiration is the chemical pathway through which carbohydrate is broken down (oxidized) into carbon dioxide and water, releasing the energy stored in the carbohydrate molecules. This is represented by the formula: Carbohydrate + O2 ���� H2O + CO2 + Energy. (The multiple arrows indicate many sequential chemical reactions.) Green plants manufacture carbohydrates by photosynthesis. Animals acquire their carbohydrates by eating plants or other animals.
Photosynthesis is somewhat the reverse of respiration: carbon dioxide and water are combined to form larger molecules of carbohydrate, with the addition of energy from sunlight: H2O + CO2 + Energy ���� Carbohydrate + O2. Water is absorbed through the roots, and CO2 diffuses into the leaves through the stomates (valved pores in leaf and stem surfaces). The plant joins several carbon dioxide molecules and adds hydrogen atoms split from water molecules to form molecules of sugar (simple carbohydrate). Surplus oxygen atoms from the water molecules are released through the stomates as oxygen gas (O2).
When you see the word �carbo-hydrate,� think �stored energy� and �calories.� Plants store energy for long-term use in the form of starch, which is a complex carbohydrate consisting of long chains of sugar molecules. When a plant needs energy to grow new leaves or flowers, it does exactly what animals do�it respires carbohydrate to release the stored energy. The complex respiratory pathway of scores of individual chemical reactions is nearly identical in all life forms: bacteria, mushrooms, saguaros, coyotes, and even in the highest life forms such as toads.
In contrast to plants, animals use fat as their main energy store; it has twice the number of calories per gram as do carbohydrate and protein. When animals in need of energy run low on the small amount of carbohydrate stored in the liver or circulating in the blood, they convert fat (or protein, if they run out of fat) into carbohydrate and then respire it.
The most common form of photosynthesis creates a 3-carbon sugar as its first stable product, so it�s called C3 photosynthesis. Other sugars with more carbon atoms are later synthesized from this first one. More than 90 percent of all plant species use C3 exclusively, but there are two specialized supplementary variations.
One variant is called C4 photosynthesis because the first stable product is a 4-carbon sugar. Plants with C4 metabolism actively transport carbon dioxide to localized bundles of photosynthetic tissue. This process offers improved efficiencies under hot, sunny conditions. C4 plants use carbon dioxide more efficiently (by bypassing photorespiration) and lose less water through transpiration (water evaporated from inside plants) per unit of carbohydrate made. The overall result is that C4 plants can grow much faster under high temperatures than most C3 plants. The majority of summer-growing grasses in warm climates are C4. So are many other summer-growing plants, especially weeds (invasive pioneer plants) that seem to spring up overnight, such as pigweed (Amaranthus spp.) and summer spurges (Euphorbia hyssopifolia and others), as well as devil�s claw (Proboscidea spp.), and many saltbushes (Atriplex spp.). Only about 3 percent of all the Earth�s plant species are known to use C4, but a number of them are vital crops, such as corn, sorghum, and sugar cane. Another variant of photosynthesis, cam, is discussed under the succulence section of this chapter on page 135. ![]()
Coping with Desert Climate
The impression that the desert environment is hostile is strictly an outsider�s viewpoint. Adaptation enables indigenous organisms not merely to survive here, but to thrive. Furthermore, specialized adaptations often result in a requirement for the seasonal drought and heat. For example, the saguaro, well adapted to its subtropical desert habitat, cannot survive in a rain forest or in any other biome, not even a cold desert. In these other places it would rot, freeze, or be shaded out by faster growing plants.
Aridity is the major�and almost the only�environmental factor that creates a desert, and it is this functional water deficit that serves as the primary limitation to which desert organisms must adapt. Desert plants survive the long rainless periods with three main adaptive strategies: succulence, drought tolerance, and drought evasion. Each of these is a different but effective suite of adaptations for prospering under conditions that would kill plants from other regions. ![]()
Succulence
As a group succulents are the most picturesque desert plants. They capture our attention because they look nothing like the familiar plants of the temperate zone where most people live. Their vernacular names suggest how they command our attention: elephant tree, boojum, jumping cholla, creeping devil, and shindagger. Spanish names translate into equally colorful terms such as dragon�s blood, child-killer, and old man�s head. Even some scientific names are inspired by the plants� characteristics: Ferocactus (as in ferocious), Opuntia molesta (the molesting-spined cactus), O. invicta (the invincible point), and Agave jaiboli (as in a highball cocktail, because liquor is made from it).
Succulent plants store water in fleshy leaves, stems or roots in compounds or cells from which it is not easily lost (see photo on page 135). All cacti are succulents, as are such non-cactus desert dwellers as agaves, aloes, elephant trees, and many euphorbias. Several other adaptations are essential for the water-storing habit to be effective. ![]()
GETTING WATER Succulents must be able to absorb large quantities of water in short periods, and they must do so under unfavorable conditions. Because roots take up water by passive diffusion, succulents can absorb water only from soil that is wetter than their own moist interiors. Desert soils seldom get this wet and don�t retain surplus moisture for long. Desert rains are often light and brief, barely wetting the top few inches (centimeters) of soil, which may dry out after just a day or two of summer heat. To cope with these conditions, nearly all succulents have extensive, shallow root systems. A giant saguaro�s root system is just beneath the soil surface and radiates as far as the plant is tall. The roots of a two-foot-tall cholla in an extremely arid site may be thirty feet (9 m) long. Most succulents, in fact, rarely have roots more than four inches (10 cm) below the surface and the water-absorbing feeder roots are mostly within the upper 1/2 inch (1.3 cm). Agaves are an exception in lacking extensive root systems; most of the roots don�t extend much beyond the spread of the leaf rosette. Instead, the leaves of these plants channel rain to the plants� bases. ![]()
CONSERVING WATER A succulent must be able to guard its water hoard in a desiccating environment and use it as efficiently as possible. The stems and leaves of most species have waxy cuticles that render them nearly waterproof when the stomates are closed. Water is further conserved by reduced surface areas; most succulents have few leaves (agaves), no leaves (most cacti), or leaves that are deciduous in dry seasons (elephant tree [Bursera spp.], boojums [Fouquieria columnaris]). The water is also bound in extracellular mucilages and inulins�compounds that hold tightly onto the water.
Many succulents possess a water-efficient variant of photosynthesis called CAM, an acronym for Crassulacean Acid Metabolism. The first word refers to the stonecrop family (Crassulaceae) in which the phenomenon was first discovered. (Dudleya is in this family, as are hen-and-chickens and jade plant.) cam plants open their stomates for gas exchange at night and store carbon dioxide in the form of an organic acid. During the day the stomates are closed and the plants are nearly completely sealed against water loss; photosynthesis is conducted using the stored carbon dioxide. At night the temperatures are lower and humidity higher than during the day, so less water is lost through transpiration. Plants using CAM lose about one-tenth as much water per unit of carbohydrate synthesized as do those using standard C3 photosynthesis. But there is a trade-off: the overall rate of photosynthesis is slower, so CAM plants grow more slowly than most C3 plants. (An additional limitation is the reduced photosynthetic surface area of most succulents compared with �ordinary� plants.)
The equilibrium between gaseous carbon dioxide and the organic acid is dependent on temperature. Acid formation (carbon dioxide storage) is favored at cool temperatures; higher temperatures stimulate release of carbon dioxide from the acid. Thus CAM works most efficiently in climates that have a large daily temperature range, such as arid lands. Cool nights allow much carbon dioxide to be stored as acid, and the warm days cause most of the carbon dioxide to be released for photosynthesis. (A note of interest: A plant in CAM mode will store enough acid to impart a sour taste in early morning; the flavor becomes bland by afternoon when the acid is used up. But don�t taste indiscriminately�many succulents are poisonous!)
Many succulents possess CAM, as do semisucculents such as some yuccas, epiphytic (growing on trees or rocks) orchids, and xerophytic (arid-adapted) bromeliads. Exceptions are stem succulents with deciduous, non-succulent leaves, such as elephant trees (Bursera spp.), limberbushes (Jatropha spp.), and desert roses (Adenium spp.). Succulents from hot, humid climates that lack substantial daily temperature fluctuations also usually do not use CAM. Some succulents, such as Agave deserti, can switch from CAM to C3 photosynthesis when water is abundant, allowing faster growth. Over five percent of all plant species spread among thirty or more plant families are known to use CAM.
Another crucial attribute of CAM plants is their idling metabolism during droughts. When CAM plants become water-stressed, the stomates remain closed both day and night and the fine (water-permeable) roots are sloughed off. The plant�s stored water is essentially sealed inside and gas exchange greatly decreases. However, a low level of respiration (oxidation of carbohydrate into water, carbon dioxide and energy) is carried out within the still-moist tissues. The carbon dioxide released by respiration is recycled into the photosynthetic pathway to make more carbohydrate, and the oxygen released by photosynthesis is recycled for respiration. Thus the plant never goes completely dormant but is metabolizing slowly�idling. (This sounds like perpetual motion, but it isn�t. The recycling isn�t 100 percent efficient, so the plant will eventually exhaust its resources.) Just as an idling engine can rev up to full speed more quickly than a cold one, an idling CAM plant can resume full growth in twenty-four to forty-eight hours after a rain. Agaves can sprout visible new roots just five hours after a rain, whereas it may take a couple of weeks for a dormant nonsucculent shrub to resume full metabolic activity. Therefore, succulents can take rapid and maximum advantage of the soil moisture from a summer rain before it quickly evaporates. The combination of shallow roots and the CAM-idling which allows rapid response enables succulents to benefit from rain even in amounts less than G inch (6 mm). ![]()
PROTECTION Stored water in an arid environment requires protection from thirsty animals. Most succulent plants are spiny, bitter, or toxic, and often all three. Some unarmed, nontoxic species are restricted to inaccessible locations. Smooth prickly pear (Opuntia phaeacantha var. laevis) and live-forever (Dudleya spp.) grow on vertical cliffs or within the canopies of armored plants. Still others rely on camouflage; Arizona night-blooming cereus (Peniocereus greggii) closely resembles the dry stems of the shrubs in which it grows.
These adaptations are all deterrents that are never completely effective. Evolution is a continuous process in which some animals develop new inheritable behaviors to avoid spines or new metabolic pathways to neutralize the toxins of certain species. In response the plants are continually improving their defenses. For example, packrats can handle even the spiniest chollas and rarely get stuck. They also eat prickly pear for water and manage to excrete the oxalates which could clog the kidneys of some other animals. Toxin-tolerant insects often incorporate their host plant�s toxins into their own tissues for protection against their predators. ![]()
Drought Tolerance
Drought-tolerant plants often appear to be dead or dying during the dry seasons. They�re just bundles of dry sticks with brown or absent foliage, reinforcing the myth that desert organisms are engaged in a perpetual struggle for survival. They�re simply waiting for rain in their own way, and are usually not suffering or dying any more than a napping dog is near death .
Drought tolerance or drought dormancy refers to desert plants� ability to withstand desiccation. A tomato plant will wilt and die within days after its soil dries out. But many nonsucculent desert plants survive months or even years with no rain. During the dry season the stems of brittlebush and bursage are so dehydrated that they can be used as kindling wood, yet they are alive. Drought-tolerant plants often shed leaves during dry periods and enter a deep dormancy analogous to torpor (a drastic lowering of metabolism) in animals. Dropping leaves reduces the surface area of the plant and thus reduces transpiration. Some plants that usually retain their leaves through droughts have resinous or waxy coatings that retard water loss (creosote bush, for example).
The roots of desert shrubs and trees are more extensive than are those of plants of the same size in wetter climates. They extend laterally two to three times the diameter of the canopy. Most also exploit the soil at greater depths than the roots of succulents. The large expanses of exposed ground between plants in deserts are usually not empty. Dig a hole almost anywhere except in active sand dunes or the most barren desert pavement and you are likely to find roots.
Rooting depth controls opportunities for growth cycles. In contrast to the succulents� shallow-rooted, rapid-response strategy, a substantial rain is required to wet the deeper root zone of shrubs and trees. A half-inch is the minimum for even the smaller shrubs �more for larger, deeper-rooted plants. It takes a couple of weeks for dormant shrubs such as brittlebush (Encelia farinosa) and creosote bush (Larrea tridentata) to produce new roots and leaves and resume full metabolic activity after a soaking rain. The tradeoff between this strategy and that of succulents is that once the deeper soil is wetted, it stays moist much longer than the surface layer; the deeper moisture sustains growth of shrubs and trees for several weeks.
Mesquite trees (Prosopis spp.) are renowned for having extremely deep roots, the champion reaching nearly 200 feet. But these riparian specimens are not drought-tolerating trees�their roots are in the water table. Most large floodplain mesquites die if the water table drops below forty feet, and mesquites growing away from waterways remain short and shrubby. No desert plant is known to use very deep roots as a primary strategy for survival. In fact, the root systems of most trees �including mesquites�are mostly confined to the upper three feet of soil. Few rains penetrate deeper than this, and at greater depths there is little oxygen to support root respiration.
In contrast to succulents that can take up water only from nearly saturated soil, drought tolerant plants can absorb water from much drier soil. A creosote bush can obtain water from soil that feels dust-dry to the touch. Similarly these plants can continue to photosynthesize with low leaf-moisture contents that would be fatal to most plants.
Some plants in this adaptive group are notoriously difficult to cultivate, especially in containers. It seems paradoxical that desert ferns and creosote bushes, among the most drought- tolerant of desert plants, can be kept alive in containers only if they are never allowed to dry out. The reason is that these plants can survive drought only if they dry out slowly and have time to make gradual physiological adjustments. If a potted plant misses a watering, the small soil volume dries out too rapidly to allow the plant to prepare for dormancy, so it dies. Researchers showed that some spike mosses (Selaginella spp.) must dehydrate over a five to seven day period. If they dry more rapidly they lack time to adjust, and if drying takes longer than a week they exhaust their energy reserves and starve to death. (Selaginella lepidophylla from the Chihuahuan Desert is widely sold as a novelty under the name �resurrection fern.� Rehydration and resumption of active life takes only a few hours.) ![]()
Drought Evasion
Interstate 40 from Barstow to Needles, California traverses some of the emptiest land in the West. It dashes as straight as it can through 130 miles (200 km) of dry valleys that are almost devoid of human settlements. The vegetation is simple, mostly widely-scattered creosote bushes. It�s difficult to tell if you�re driving through the Mohave or Sonoran desert. The small, rocky mountain ranges interrupting the valleys beckon to true desert lovers, but the drive is just plain bleak to most folks. The exits on this freeway average ten miles (16 km) apart and connect to two-lane roads that shoot straight over the distant horizon with no visible destinations. You rarely see a vehicle on any of them.
Frequent travelers on this freeway become accustomed to its monotony until they think they know what to expect. The creosote bush may turn green if there�s been a rain; ocotillo always flowers in April; most of the time it�s just brown gravel and brown bushes. Then one spring travelers were astonished to discover the ground between the bushes literally carpeted with flowers. It happened in March 1998, when for three weeks the freeway bisected a nearly unbroken blanket of desert sunflowers forty miles long and ten miles wide. At every exit-to-nowhere several cars and trucks were pulled off and people wandered through the two-foot-deep sea of yellow. Those with long memories may have recalled that the same thing happened in 1978. Perhaps they wondered where these flowers came from, and where they were during the intervening twenty years.
Those desert sunflowers (Geraea canescens) were annual wildflowers, plants that escape unfavorable conditions by �not existing� during such periods. Annuals complete their life cycles during brief wet seasons, then die after channeling all of their life energy into producing seeds. Seeds are dormant propagules with almost no metabolism and great resistance to environmental extremes. (A propagule is any part of a plant that can separate from the parent and grow into a new plant, for example, a seed, an agave aerial plantlet, a cholla joint.) Seeds wait out adverse environmental conditions, sometimes for decades, and will germinate and grow only when specific requirements are met.
Wildflower spectacles like the one described above are rare events. Mass germination and prolific growth depend on rains that are both earlier and more plentiful than normal. The dazzling displays featured in photographic journals and on postcards occur about once a decade in a given place. In the six decades between 1940 and 1998 there have been only four documented drop-everything-and-go-see-it displays in southern Arizona: 1941, 1978, 1979, and 1998. During that period only the displays of 1978 and 1998 were widespread throughout both the Sonoran and Mohave deserts.
Annuals in the Sonoran Desert can be divided into three groups, based on time of germination and flowering. Winter-spring species are by far the most numerous. The showy wildflowers that attract human attention will germinate only during a narrow window of opportunity in the fall or winter, after summer heat has waned and before winter cold arrives. In most of the Sonoran Desert this temperature window seems to occur between early October and early December for most species. During this window there must be a soaking rain of at least one inch (2.5 cm) to induce mass germination. This combination of requirements is survival insurance: an inch of rain in the mild weather of fall will provide enough soil moisture that the resulting seedlings will probably mature and produce seeds even if almost no more rain falls in that season. (Remember that one of the characteristics of deserts is low and undependable rainfall.) If the subsequent rainfall is sparse, the plants remain small and may produce only a single flower and a few seeds, but this is enough to ensure a future generation . There is still further insurance: even under the best conditions not all of the seeds in the soil will germinate; some remain dormant. For example, a percentage of any year�s crop of desert lupine seeds will not germinate until they are ten years old. The mechanisms that regulate this delayed germination are not well understood.
The seedlings produce rosettes of leaves during the mild fall weather, grow more slowly through the winter (staying warm in the daytime by remaining flat against the ground), and bolt into flower in the spring. Since the plants are inconspicuous until they begin the spring bolt, many people mistakenly think that spring rains produce desert wildflower displays.
There is a smaller group of annual species that grow only in response to summer rains. Annual devil�s claw (Proboscidea parviflora) and Arizona poppy (Kallstroemia grandiflora) are among the few showy ones.
A third group consists of a few opportunistic species which will germinate in response to rain at almost any season. Most of these lack showy flowers and are known only to botanists, but desert marigold (Baileya multiradiata) is a conspicuous exception; it is actually not an annual, but rather a short-lived perennial in most of its range. A few species of buckwheats (Eriogonum) germinate in fall or winter and flower the following summer.
The annual habit is a very successful strategy for warm-arid climates. There are no annual plants in the polar regions or the wet tropics. In the polar zones the growing season is too short to complete a life cycle. In both habitats the intense competition for suitable growing sites favors longevity. (Once you�ve got it, you should hang onto it.) Annuals become common only in communities that have dry seasons, where the perennials are widely spaced because they must command a large soil area to survive the drier years. In the occasional wetter years, both open space and moisture are available to be exploited by plants that can do so rapidly. The more arid the habitat, the greater the proportion of annual species in North America. (The percentage decreases in the extremely arid parts of the Saharan-Arabian region.) Half of the Sonoran Desert�s flora is comprised of annual species. In the driest habitats, such as the sandy flats near Yuma, Arizona, up to ninety percent of the plants are annuals.
Winter annuals provide most of the color for our famous wildflower shows. Woody perennials and succulents can be individually beautiful, but their adaptive strategies require them to be widely-spaced, so they usually don�t create masses of color. A couple of exceptions are brittlebush when it occurs in pure stands, and exten sive woodlands of foothill palo verde (Cercidium microphyllum). The most common of the showy winter annuals that contribute to these displays in southern Arizona are Mexican gold poppy (Eschscholtzia mexicana), lupine (Lupinus sparsiflorus), and owl-clover (Castilleja exserta, formerly Orthocarpus purpurascens).
One of the contributing factors to the great number of annual species is niche separation. (A niche is an organism�s ecological role; for example, sand verbena is a butterfly-pollinated winter annual of sandy soils.) Most species have definite preferences for particular soil textures, and perhaps soil chemistry as well. For example, in the Pinacate region of northwestern Sonora there are places where gravels of volcanic cinder are dissected by drainage channels or wind deposits of fine silt. In wet years Nama demissum (purple mat) grows abundantly on the gravel and the related Nama hispidum (sand bells) on the silt. I have seen the two species within inches of each other where these soil types meet, but not one plant of either species could be found on the other soil. There are specialists in loose sand such as dune evening primrose (Oenothera deltoides) and sand verbena (Abronia villosa), and others are restricted to rocky soils, such as most caterpillar weeds (Phacelia spp.). This phenomenon of occupying different physical locations is spatial niche separation.
Another diversity-promoting phenomenon is temporal niche separation: the mix of species at the same location changes from year to year. Seeds of the various species have different germination requirements. The time of the season (which influences temperature) and quantity of the first germination-triggering rain determines which species will dominate, or even be present at all in that year. Of the three most common annuals of southern Arizona listed above, any one may occur in a nearly pure stand on a given hillside in different years, and occasionally all three are nearly equally abundant. This interpretation of the cause of these year-to-year variations is a hypothesis based on decades of empirical observation. Much more research is needed to discover the ecological requirements of most species of desert annuals. And of course the Sonoran Desert�s two rainy seasons provide two major temporal niches. Summer and winter annuals almost never overlap.
The dramatic wildflower shows are only a small part of the ecological story of desert annuals. For each conspicuous species there are dozens of others that either have less colorful flowers or don�t grow in large numbers. Every time the desert has a wet fall or winter it will turn green with annuals, but it will not always be ablaze with other colors. One of the most common winter annuals is desert plantain (Plantago insularis). It usually grows only a few inches tall and bears spikes of tiny greenish flowers, but billions of plants cover many square miles in good years. The tiny seeds are covered with a soluble fiber which forms a sticky mucilage when wet by rain; this aids germination by retaining water around the seed and sticking it to the ground. A related species from India is the commercial source of psyllium fiber (Metamucil� for example). The buckwheat family (Polygonaceae) is well- represented. There are more than a score of skeleton weeds (Eriogonum spp.) and half as many spiny buckwheats (Chorizanthe spp.), most of which go unnoticed except by botanists (see species accounts). Fiddlenecks (Amsinckia spp., Boraginaceae) may grow in solid masses over many acres, but the tiny yellow flowers don�t significantly modify the dominant green of the foliage. These more modest species produce more biomass than the showy wildflowers in most years, and thus form the foundation of a great food pyramid.
Some perennials also evade drought much as annuals do, by having underground parts that send up stems, leaves, and flowers only during wet years. Coyote gourd (Cucurbita digitata) and perennial devil�s claw (Proboscidea althaeifolia) have fleshy roots that remain dormant in dry years. Desert larkspur (Delphinium parryi) is a perennial that has woody rootstocks but also sprouts only in wetter years. Desert mariposa (Calochortus kennedyi) and desert lily (Hesperocallis undulata) have bulbs that may remain dormant for several years until a deep soaking rain awakens them.
Our desert wildflower displays are in jeopardy from invasive exotic plants. Species such as Russian thistle (Salsola tragus, also called S. kali), mustards (especially Brassica tournefortii), filaree (Erodium cicutarium), and Lehmann�s lovegrass (Eragrostis lehmannii) are more aggressive than most of the native annuals and are crowding them out in many areas where they have become established. Some are still increasing their geographic ranges with every wet winter. Disturbed sites such as sand dunes, washes (naturally disturbed by wind and water, respectively), roadsides, and livestock-grazed lands are particularly vulnerable to invasion by these aliens. ![]()
Combined Drought Adaptations
These three basic drought-coping strategies�succulence, drought tolerance, and drought avoidance�are not exclusive categories. Ocotillo behaves as if it were a CAM-succulent, drought deciduous shrub, but it is neither CAM nor succulent (see details in the species accounts). The genus Portulaca contains species that are succulent annuals. The seeds may wait for a wet spell to germinate, but the resulting plants can tolerate a moderate drought. The semisucculent yuccas have some water storage capacity, but rely on deep roots to obtain most of their water. Mesquite trees are often phreatophytes (plants with their roots in the water table), but some species can also grow as stunted shrubs on drier sites where ground water is beyond their reach. ![]()
Adaptations to Other Desert Conditions
Water scarcity is the most important�but not the only�environmental challenge to desert organisms. The aridity allows the sun to shine unfiltered through the clear atmosphere continuously from sunrise to sunset. This intense solar radiation produces very high summer temperatures which are lethal to nonadapted plants. At night much of the accumulated heat radiates through the same clear atmosphere and the temperature drops dramatically. Daily fluctuations of 40�F (22�C) are not uncommon when the humidity is very low.
Microphylly (the trait of having small leaves) is primarily an adaptation to avoid overheating; it also reduces water loss. A broader surface has a deeper boundary layer of stagnant air at its surface, which impedes convective heat exchange. A leaf up to 1/2 inch (10 mm) across can stay below the lethal tissue temperature of about 115�F (46�C) on a calm day with its stomates closed. A larger leaf requires transpiration through open stomates for evaporative cooling. Since the hottest time of year is also the driest, water is not available for transpiration. Non- succulent large-leafed plants in the desert environment would overheat and be killed. Desert gardeners know that tomatoes will burn in full desert sun even if well watered; their leaves are just too big to stay cool. Desert plants that do have large leaves produce them only during the cool or rainy season or else live in shaded microhabitats. There are a few mysterious exceptions, such as jimson weed (Datura wrightii) and desert milkweed (Asclepias erosa). Perhaps their large tuberous roots provide enough water for transpiration even when the soil is dry.
Leaf or stem color, orientation, and self-shading are still more ways to adapt to intense light and heat. Desert foliage comes in many shades, but rarely in typical leaf-green. More often leaves are gray-green, blue-green, gray, or even white. The light color is usually due to a dense covering of trichomes (hairlike scales), but is sometimes from a waxy secretion on the leaf or stem surface. Lighter colors reflect more light (= heat) and thus remain cooler than dark green leaves. Brittlebush and white bursage leaves show no green through their trichomes during the dry season, while desert agave (Agave deserti) is light gray due to its thick, waxy cuticle. Other plants have leaves or stems with vertical orientations; two common examples are jojoba and prickly pear cactus. This orientation results in the photosynthetic surface facing the sun most directly in morning and late afternoon. Photosynthesis is more efficient during these cooler times of day. Prickly pear pads will burn in summer if their flat surfaces face upward. Some cacti create their own shade with a dense armament of spines; teddy bear cholla (Opuntia bigelovii) is one of the most striking examples. ![]()
Pollination Ecology and Seed Dispersal of Desert Plants
(this portion of the chapter is also available in Spanish: Ecología de la polinización y dispersión de las semillas de las plantas del desierto)
Flowers are very useful for identifying plants and providing aesthetic pleasure for humans, but they have a more vital function�they are the sexual reproductive organs of plants. Many plants also have methods of asexual (vegetative) reproduction, which produces offspring that are genetically identical to the parent: root-sprouting (limberbush, palo verde, aspen), stolons and rhizomes (agaves, strawberries, many grasses), and aerial plantlets (some agaves, mother-of-millions, kalanchoe). All of the progeny of asexual reproduction are clones of their parent plants. (A clone is a group of organisms that are genetically identical; in the case of flowering plants each clone originates from a single seed.) Horticulturists have developed additional methods of plant cloning that are valuable in perpetuating superior varieties of plants: cutting, grafting, and tissue culture. The �Kadota� fig is a cultivar (contraction for cultivated variety) that has been propagated by cuttings for at least two millennia; it is described under a different name in the writings of Pliny the Younger.
In contrast, sexual reproduction combines half the genes from each of two parents, so sexually produced offspring are different from either of their parents and from one another. This variation is the raw material of natural selection which in turn results in evolution. A species that cannot reproduce sexually�there are quite a few among both plants and animals�is at greater risk of extinction if its environment changes, because it cannot adapt to new conditions.
Pollination is the transfer of pollen from an anther onto the stigma of a flower. The pollen then grows a tube that penetrates the style down to the ovary; sperm cells swim down the tube and fertilize the ova. Fertilized ova develop into seeds, which are the sexual propagules of flowering plants.
Outcrossing (pollination by pollen from another plant) is evolutionarily advantageous because the offspring are more variable than those from self- pollination. Variability increases plants� probability of surviving in an ever-changing environment. (But self- pollination is still sexual reproduction which results in different combinations of genes and therefore allows evolutionary change, as vegetative cloning does not.) Plants have many adaptations that increase the likelihood of outcrossing.
Because plants are rooted in the ground and can�t get together to mate, they must employ an agent to transport pollen between plants. From this need widespread and complex kinds of mutualism (mutually beneficial interactions) have evolved between plants and animals. The pollen-transporting agent is frequently an insect or other flying animal. (Flying animals are more mobile than grounded species, and thus more likely to visit widely-separated plants.) In order to get pollinated, a flower must both make its presence known (advertise), and provide an incentive (a reward) for an animal to make repeated visits to flowers of the same species. The advertisements are fragrance and/or conspicuous color. Two kinds of food are the usual reward. Nectar is a sugar solution that provides energy for flight. Flying requires much more energy than terrestrial locomotion. Pollen, besides being the male gene-bearer of a flower, is also rich in proteins essential for maintaining animal tissues and for raising young. In place of nectar some flowers offer oil (fat), another energy food. Others provide fragrances that the pollinator gathers to use for its own reproductive advertisement, and a few fascinating species employ deceit and provide no reward (see the species account on pipevine for an example).
The sugar in nectar and the protein in pollen are expensive to produce, so there is selective pressure to use these resources efficiently. It is important that animals other than the pollinators do not eat (steal) the nectar and pollen, and that the pollinators transport pollen to other flowers of the same species and deposit it in the right place. Natural selection has produced specialization: most plants with animal-pollinated flowers attract only a few species of animals which have the right size and behavior to reach the reward and pick up pollen. The more than 100 million years of coevolution between flowering plants and their pollinators has greatly contributed to the huge number of species in both kingdoms (300,000 flowering plants, 350 hummingbirds, and 15,000 known bees in the world). It also explains why there are so many different shapes and colors of flowers.
Flowers can be classified into several pollination syndromes according to their pollinators. (A syndrome is a set of characteristics associated with a specific phenomenon.) This is not the same classification as systematic taxonomy and does not reflect the evolutionary relationships among plants. Species in the same family or even the same genus may attract different pollinators.
The hummingbird pollination syndrome is one of the most easily recognized. Hummingbirds are large compared to most insects, almost unique in their ability to feed while hovering, and daytime-active; they have no sense of smell, but have long narrow beaks and tongues that can probe deep narrow tubes, and excellent color vision. Hummingbird flowers tend to be long-tubular, non-fragrant, sideways- or downward-facing, day-blooming, and brightly colored. Bees and most other animals cannot easily land on a hanging flower, and even if they succeed they cannot reach the nectar at the base of the narrow tube.
There are common misconceptions that all hummingbird flowers are red and that hummingbirds can see only the warm colors of the spectrum. It is true that most hummingbird flowers in the temperate biomes are red, but in the tropics they come in many colors. The predominance of red in temperate hummingbird flowers may be a disincentive to bees. Bees are aggressive pollen collectors in temperate climates. But they cannot see red, so red flowers do not appear conspicuous to them.
Wind-pollinated plants make no investment in attracting animals; their flowers lack fragrance or showy parts. Many people would not recognize them as flowers at all. Prodigious quantities of pollen are released, an infinitesimal proportion of which lands on a receptive stigma of the same species. While this seems inefficient, it is obviously effective, judging from the successful groups of plants with this syndrome. Conifers, most riparian trees (such as willows and sycamores), oaks, and grasses are all wind-pollinated. Conifers and grasses are the dominant plants in the two biomes that bear their names. Grasses occur in most biomes and comprise the sixth largest family of plants with about 9000 species worldwide. Wind pollination is not always entirely passive (see the species account for jojoba). ![]()
Seed Dispersal
Seeds generally need to be transported some distance from the parent plant in order to find a suitable site for establishment. Some plants have wind-dispersed seeds, which are occasionally blown many miles from their origins. This means of dispersal is common among pioneer plants (plants that are adapted to colonizing disturbed habitats). Because of their superior ability to invade newly-disturbed ground, pioneer plants comprise many of our agricultural and garden weeds. Moreover, most annual crops are domesticated pioneer plants. That�s why we need to plow (disturb) fields in order to grow them.
Many plants use animals to disperse their seeds in another complex coevolutionary process. Small, brightly-colored fruits such as hackberry and boxthorn are offered as food for birds that swallow them whole. Other fruits such as those of hedgehog cacti are large and birds feed on them repeatedly. Some bird fruits are sticky, such as mistletoe berries; a few stick to the bird�s bill until wiped off on a branch while others are successfully swallowed. The seeds of bird fruits are typically small and hard; they pass through birds� guts undamaged and may be deposited many miles from the parent plant.
Mammal-dispersed fruits tend to be larger, aromatic, not colorful (most non-primate mammals have poor color vision), and usually have larger seeds than bird fruits. The animal often transports the fruits a short distance (compared to the flying distances of many birds) to a safer place before eating the pulp and dropping at least some of the seeds. The seeds of coyote gourds (Cucurbita spp.) may be dispersed in this manner. Coyotes swallow the whole fruits of palm trees; they digest the thin pulp and excrete the hard seeds intact. Since seeds contain energy stores to nourish the germinating embryo, seeds themselves are also nutritious food for mammals and birds. Some plants offer their seeds without juicy pulp to attract mammals. Pocket mice and antelope squirrels gather the abundant seeds of foothill palo verdes and bury them as food caches for the dry season. The animals don�t eat all that they bury, so some seeds remain in the ground and germinate when the rains come. (Birds that specialize in eating seeds, as opposed to fruits containing seeds, crush and digest the seeds and therefore do not disperse viable propagules.)
Even in the desert some seeds are water-dispersed. Blue palo verde (Cercidium floridum) grows mostly along washes. Flash floods disperse the very hard, waterproof seeds downstream, scarifying (abrading the surface of) them in the process. In the absence of scarification these seeds must weather in the ground for a few years before the seed coats become permeable and permit germination.
The timing of seed maturation is crucial for many plants. The less time seeds are present before they sprout, the greater is their chance of survival. The tropically derived plants in our region germinate with the summer rains. These species usually flower in spring and their fruits ripen shortly before the arrival of the summer rainy season. Palo verde and saguaro are examples. Other plants produce large quantities of seeds and rely on camouflage or burial in the soil to conceal some of them from hungry animals. Brittlebush, for example, flowers and seeds in spring, but the seeds germinate with fall rains. Annuals do the same. ![]()
Flowering Seasons in the Sonoran Desert
The Spring Flowering Season
The spring flowering season in the Arizona Upland subdivision spans from mid February to mid June with a peak from mid March to late April depending on rainfall and temperatures during the growing season. In the warmest areas of the Lower Colorado River Valley subdivision it is normally a couple of weeks earlier, though it sometimes starts as early as November. The different life forms which dominate at different times vary in their showyness and reliability. The early-blooming winter annuals can create an incredible display, but do so only rarely. Later-blooming species bloom more dependably, but mostly not in great masses of color. The progression of spring bloom described below is for average years near Tucson. It may be three weeks earlier or later depending on weather, elevation, and latitude.
WINTER ANNUALS such as poppy (Eschscholtzia mexicana), lupines (Lupinus sparsiflorus and others), and owl-clover (Castilleja exserta) create the vast carpets of color for which the Sonoran and Mohave deserts are so famous. This event may occur between late February and mid April, usually in mid March. Annuals are highly dependent on rainfall. Massive and widespread displays occur only about once a decade, when the winter rainy season is both earlier and wetter than normal. Good shows happen in localized areas every three or four years. A good bloom cannot be reliably predicted more than a week or two before it begins, and usually lasts at peak beauty for only two weeks. Seeing such a bloom requires being able to travel on short notice, and perhaps great distances. Death Valley may be spectacular in a year when Organ Pipe Cactus National Monument is poor. The high Mohave Desert may peak two or three weeks later than the lower-elevation and more southerly Sonoran Desert. A good bloom may occur in a remote area and remain undiscovered.
HERBACEOUS PERENNIALS AND SMALL SHRUBS  such as penstemon (especially Penstemon parryi, shown here), brittlebush (Encelia farinosa), and fairy duster (Calliandra eriophylla) also require rain to bloom but are less sensitive to its timing. They are somewhat more dependable than the annuals, making a good show in about half of the years and peaking some time in March. These species usually grow as individuals or in small patches and do not create masses of color.
such as penstemon (especially Penstemon parryi, shown here), brittlebush (Encelia farinosa), and fairy duster (Calliandra eriophylla) also require rain to bloom but are less sensitive to its timing. They are somewhat more dependable than the annuals, making a good show in about half of the years and peaking some time in March. These species usually grow as individuals or in small patches and do not create masses of color.
CACTI, because they store water, are fairly independent of rain. They bloom well nearly every year though wetter years produce more flowers. The greatest diversity of spring-blooming species can be seen in April. The cactus show continues as the abundant prickly pears bloom in early May, followed by saguaros from mid May to mid June.
 TREES AND LARGE SHRUBS are fairly dependable bloomers, though flowers will be sparse in dry years. Creosote bush (Larrea tridentata) and whitethorn acacia (Acacia constricta, shown here) both bloom mainly in spring and sometimes again in summer. Blue palo verde (Cercidium floridum) turns bright yellow in late April, followed two weeks later by the much more abundant but paler yellow foothills palo verde (C. microphyllum). Desert ironwood trees (Olneya tesota) bloom heavily about every other year with masses of lavender flowers, usually in late May. The abundant ocotillo reliably produces spikes of red flowers throughout April. These species bloom about two weeks earlier in western Arizona.
TREES AND LARGE SHRUBS are fairly dependable bloomers, though flowers will be sparse in dry years. Creosote bush (Larrea tridentata) and whitethorn acacia (Acacia constricta, shown here) both bloom mainly in spring and sometimes again in summer. Blue palo verde (Cercidium floridum) turns bright yellow in late April, followed two weeks later by the much more abundant but paler yellow foothills palo verde (C. microphyllum). Desert ironwood trees (Olneya tesota) bloom heavily about every other year with masses of lavender flowers, usually in late May. The abundant ocotillo reliably produces spikes of red flowers throughout April. These species bloom about two weeks earlier in western Arizona.
SUMMARY
If you want to see the famous carpets of color, keep abreast of local news from Palm Springs to Tucson and from Death Valley to northern Mexico. Begin checking in late February and be ready to travel on short notice. You�ll find masses of annuals somewhere in this area about once every three or four years. If you want dependability and will settle for less quantity, success is almost guaranteed in the middle half of April. ![]()
The Summer Flowering Season
This season begins a few weeks after the first summer rain and continues into late fall. Though there are many beautiful species to be seen, there are rarely massive displays of color in this season, because the summer rains are more sporadic and localized than the winter rains and the soil dries rapidly in the heat.
SUMMER ANNUALS such as summer poppy (Kallstroemia grandiflora) and devil�s claw (Proboscidea parviflora) germinate within a few days after the first soaking summer rain and begin to flower as soon as three weeks later. Chinchweed (Pectis papposa) is the most widely-adapted summer annual; it ranges from New Mexico into the central Mohave Desert where it is the only summer annual (summer rains are uncommon in the Mohave). It can form showy carpets of yellow when rains are abundant.
 HERBACEOUS PERENNIALS AND SMALL SHRUBS bloom opportunistically if they get enough rain. Trailing four-o�clock (Allionia incarnata) and desert marigold (Baileya multiradiata) are nonseasonal, flowering in response to rain in all but the coldest months. Fairy duster will also bloom again in wet summers, but not as profusely as in spring. Sacred datura (Datura wrightii) is mainly a summer perennial though it may begin flowering as early as April in warmer areas. There are several woody shrubs that bloom in late fall. Most are composites such as turpentine bush (Isocoma tenuisecta) and desert broom (Baccharis sarothroides). Desert senna (Senna covesii) and Coulter hibiscus (Hibiscus coulteri) flower in response to any warm rain and peak in summer when most such rain occurs. Desert zinnia (Zinnia pumila) is truly biseasonal, flowering well in both rainy seasons.
HERBACEOUS PERENNIALS AND SMALL SHRUBS bloom opportunistically if they get enough rain. Trailing four-o�clock (Allionia incarnata) and desert marigold (Baileya multiradiata) are nonseasonal, flowering in response to rain in all but the coldest months. Fairy duster will also bloom again in wet summers, but not as profusely as in spring. Sacred datura (Datura wrightii) is mainly a summer perennial though it may begin flowering as early as April in warmer areas. There are several woody shrubs that bloom in late fall. Most are composites such as turpentine bush (Isocoma tenuisecta) and desert broom (Baccharis sarothroides). Desert senna (Senna covesii) and Coulter hibiscus (Hibiscus coulteri) flower in response to any warm rain and peak in summer when most such rain occurs. Desert zinnia (Zinnia pumila) is truly biseasonal, flowering well in both rainy seasons.
CACTI include several summer-flowering species. The pincushion cactus Mammillaria grahamii makes buds during its previous growing season, then goes dormant during the dry season. The buds burst into bloom five days after each of the first two or three summer rains. The fishhook barrel cactus, Ferocactus wislizeni, is much larger than the pincushion and less dependent on rain; it flowers throughout August and September.
TREES AND LARGE SHRUBS are nearly all spring bloomers, but a few bloom again in summer if rains are generous. Whitethorn acacia (Acacia constricta) and velvet mesquite (Prosopis velutina) flower heavily in spring and often again in summer. Desert willow (Chilopsis linearis) flowers from spring through fall if it has enough water.
SUMMARY
Though the Sonoran Desert has two flowering peaks, there is almost always something in bloom. The only exceptions are after a hard winter freeze or during severe droughts. ![]()
Predicting Wildflower Blooms
Desert annual wildflower blooms are nearly impossible to predict for two reasons. First, the necessary conditions are not precisely known. Second, many interacting variables affect the phenomenon. Here is what we do know:
-
Spring-blooming annuals must germinate in the autumn. This is a crucial fact that most people don�t know. The �critical window� is probably between late September and early December, but differs with different species. The controlling environmental factor is temperature.
-
A �triggering rain� of at least one inch must occur during this autumn window, the earlier the better after summer heat has waned. Rains at other times will seldom trigger germination of the showy-flowered species.
-
The triggering rain must be followed by regular rains totaling at least an inch per month through March, a season total of at least five inches�seven or more are better.
In short, a really good wildflower bloom requires both an unusually early and an unusually wet winter rainy season. The rains must also be well spaced. Spectacular, widespread shows occur about once in ten years in the Sonoran and Mohave deserts. Good or better displays occur in localized areas perhaps every three or four years; these may be in remote regions and go unnoticed.
However, even when all the above conditions are met, the bloom may be mediocre or poor. And occasionally a good bloom occurs when the above conditions appear not to have been met. The latter can happen when an unusually warm rain triggers germination in winter, but the short growing season usually precludes a really good show. Factors which are suspected of preventing a show include:
-
a few weeks of warm, windy weather; the water stress triggers premature flowering;
-
a cold winter that retards growth of the seedlings;
-
high population levels of herbivores: rodents, rabbits, quail, or insects; and
-
a wet preceding summer, resulting in thick growth of summer vegetation, which in turn prevents germination of winter annuals.
When it does happen, the peak typically lasts only two weeks at a given location, sometime between late February and mid April. Most often it happens in early to mid March.
Furthermore, only certain areas ever have mass displays; soil type and vegetation cover are important factors. The rocky and densely-vegetated Tucson Mountains rarely if ever have mass blooms, whereas Picacho Peak and the Tohono O�odham (Papago) Indian Reservation do fairly regularly.
The above information pertains only to annual wildflowers such as poppies, lupines, and owl-clover. Perennials are less fussy about the timing of rainfall. Thus a late but wet rainy season can still produce good blooms of penstemon, larkspur, brittlebush, and other perennials. Some plants such as palo verdes, ocotillo, and most cacti flower nearly every year regardless of rainfall. (See other page for details.) But it is the annuals alone that produce the desert�s famous carpets of color. ![]()
The Invisible Larder
I conducted a wildlife survey in the Lower Colorado River Valley in the 1970s. The site had received almost no biologically effective rainfall for three years. Creosote bushes were almost the only plants present; they were widely-spaced and had shed most of their leaves. Yet in the kilometer (6/10 mile) long by fifty meter (150 foot) wide transect I trapped one pocket mouse overnight, and in the morning observed a whiptail lizard, a rock wren, and two black-throated sparrows. These are all resident species; not transitory migrants. What were they living on?
A persistent, large soil seed bank is an extremely important resource in arid habitats. It provides an unseen (by humans) food source for desert animals as well as survival insurance for plant species. The greater density of seed-eating animals and the abundance of decomposing microbes in the moist soils of wetter regions greatly shortens the viability of seeds. In deserts viable�and nutritious�seeds persist in large numbers through decades of drought. After a wet year there may be 200,000 seeds per square meter (square yard) of soil. Even after several dry years with little or no additional seed production there are still several thousand seeds per square meter, enough to sustain low populations of seed-eaters such as harvester ants, kangaroo rats, and sparrows. The whiptail was foraging for insects that fed on the seeds or plant detritus (partially decomposed organic matter) in the soil. As the statistician in the movie �Jurassic Park� said, �Life will find a way.� ![]()
Additional Readings
Bowers, Janice E. A full life in a small place and other essays from a desert garden. Tucson, University of Arizona Press, 1993.
Buchmann, Steven L. and Gary P. Nabhan. The Forgotten Pollinators. Washington, D.C.: Island Press, 1996.
Dykinga, Jack W. and Charles Bowden. The Sonoran Desert. NY: Harry N. Abrams, 1992.
Hanson, Roseann Beggy and Jonathan Hanson. Southern Arizona Nature Almanac. Boulder: Pruett Publishing Co, 1996.
Hartmann, William K. Desert Heart: Chronicles of the Sonoran Desert. Tucson: Fisher Books, 1989.
Imes, Rick. 1990. The practical botanist: An essential field guide to studying, classifying, and collecting plants. New York: Fireside Books/Simon and Schuster, 1990.
Larson, Gary. There�s a Hair in My Dirt! A Worm�s Story. New York: Harper-Collins, 1998.*
Nabhan, Gary P. Gathering the Desert. Tucson: University of Arizona Press, 1985.
���. The desert smells like rain. San Francisco: North Point Press, 1982.
Seuss, Dr. The Lorax. New York: Random House, 1971.* ![]()
* Though these two books are found in children�s literature, they convey the essence of ecology better than any scientific treatise I have encountered.

