






Desert Grasses
Thomas R. Van Devender and Mark A. Dimmitt
The grass family Poaceae (also known as Gramineae) has some 10,000 species and 650 to 900 genera. Only the sunflower (Asteraceae, or Compositae), legume (Fabaceae, or Leguminosae), and orchid (Orchidaceae) families are larger. The grass family has more individual plants and a wider environmental range than does any other family, reaching the limits of vegetation in polar regions and on mountaintops, enduring extremes of cold, heat, and drought, and dominating various landscapes worldwide. Grasses are the most successful monocots (seed-bearing plants with single seed leaves), and the most beneficial plants for humankind, providing highly nutritional grains and livestock forage, and preventing soil erosion. Of the 5 crops that provide almost two-thirds of the food we eat, including corn, barley, potatoes, rice, and wheat, only potatoes are not a grain.
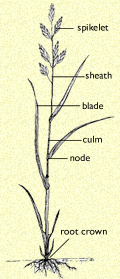
The structure of California brome
Grasses are highly specialized monocots distinguished by certain characteristics of the stems, leaves, and inflorescences. The jointed stems (culms) are round or flattened (never triangular); they are usually hollow except at the nodes (points on the stem from which leaves arise), where they are solid. The leaves generally consist of 2 main parts: a tubular sheath around the plant?s stem and a blade (the broader, expanded part). Leaves are usually single at each node, but are 2-ranked, that is, spaced alternatively on opposite sides of the stems. Very small, individual, simple flowers (florets) are grouped in inflorescences called spikelets, which are subtended by small leaf-like bracts, the glumes. The florets are enclosed by other specialized bracts (the lemma and palea).
There are grasses that use both C3 and C4 photosynthetic pathways to fix carbon into tissue (see pages 132-134). Primitive grasses such as the bamboos and modern grasses that live at higher latitudes or elevations where temperatures in the growing season are relatively cool, mostly have the more typical C3 photosynthesis predominating. However, in the intense sunlight of warmer habitats at lower latitudes and elevations, grasses with the more specialized C4 metabolism are better able to fix carbon. C4 grasses are important in tropical forests, semiarid grasslands, and warm deserts.
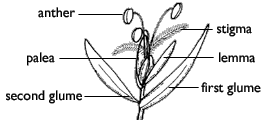
California brome floret
The earliest grasses lived in shady tropical forests. The evolution and spread of grasses undoubtedly resulted from their ability to adapt to seasonally dry habitats created as tropical deciduous forests developed in the Eocene (58 to 34 mya, million years ago). Considering their importance and taxonomic diversity, grasses have a relatively poor fossil record. While the earliest potential fossil grass pollen was described from late Cretaceous sediments, the oldest reliable megafossil grass fossils were spikelets and inflorescences from the latest Paleocene (about 58 mya). These were primitive proto-bamboos with broad leaves, quite unlike the narrowleaf modern grasses of desert grasslands and deserts. Although the early fossils could not be assigned to living grasses, their morphological details reflect the early evolution of wind pollination in a seasonally dry tropical environment. In the late Oligocene (about 30 to 24 mya) fossils of central North America, more diverse grass fossils were found, including both archaic forms and quite a few living genera. By the Miocene (24 mya), many more modern genera appeared in the fossil record. A great deal of the history of grasses was clearly not captured in the fossil record, notably the evolutionary radiation from primitive proto- bamboos to modern grasses in the Eocene and Oligocene. By the early Miocene, however, grasses in all our modern subfamilies were present, indicating that our modern taxonomic and physiologic diversity had been well established by that time.
Desert Grasses
The plants that are most conspicuous in Sonoran desertscrub communities are long-lived trees, such as foothills palo verde (Cercidium microphyllum), and cacti, such as the saguaro (Carnegiea gigantea), with grasses usually not so noticeable. Yet grasses are surprisingly numerous and play important ecological roles in desert communities.
Growth Forms
Many plants lose parts in times of stress. In temperate and tropical forests, deciduous trees drop their leaves once a year in response to cold or drought. Ocotillo (Fouquieria splendens) loses its leaves with drought, but leafs out several times a year depending on rainfall. Desert trees and shrubs such as creosote bush (Larrea tridentata), foothills palo verde, triangleleaf bursage (Ambrosia deltoidea), and many others, actually lose branches during drought or frost, becoming smaller plants; they could be considered stem- or branch-deciduous. Perhaps the ultimate type of deciduousness is that of the herbaceous perennials, including the bunch grasses of semiarid grasslands and deserts. (Perennials are plants that grow and reproduce for many years; herbaceous plants, unlike shrubs and trees, do not become woody.) With drought, frost, fire, and grazing, these plants die back to perennial crowns or roots?they are almost ?whole plant deciduous.?
When suitable moisture and temperature conditions return, bunch grasses must grow stems and photosynthetic leaf blades before flowering, and their new stems and leaves are pushed up from basal meristems. (Meristems are growing points that persist from year to year; these are basal?that is, at or below ground level?in many grasses.) This is in sharp contrast to nearly all other plants, in which the meristems are at the upper or outer stem tips of the plant and leaf-bearing stems are laid down beneath them. The advantage of this basal meristematic growth form is that when the above-ground plant parts are eaten by grazing animals or burned, the stems and leaves can be replaced rapidly, using energy stored in the crown and roots. Perennial desert grasses range in size from low turf grasses like curly mesquite grass (Hilaria belangeri) and false grama (Cathestecum brevifolium) to the nearly 3-foot (0.9 m) tall Arizona cottontop (Digitaria californica) and tanglehead (Heteropogon contortus).
There are two distinct alternative growth forms in desert grasses that differ from that of bunch grasses. A few larger grasses, notably big galleta (Pleuraphis rigida, formerly Hilaria rigida), bush muhly (Muhlenbergia porteri), and the introduced buffelgrass (Cenchrus ciliaris), are functionally more like shrubs than they are like bunch grasses. During normal dry and cool periods, their leaves die back to nodes along the stems. This gives shrubby grasses an advantage: water can move from the soil to the living nodes through the intact stems after rains; new leaves and then flowers form rapidly. With fire or severe frost, the plants die back to the crown, essentially reverting to a bunch-grass life form; massive roots allow for rapid regrowth. Bush muhly is a widespread perennial that apparently has declined dramatically with cattle grazing. Repeat photography in the Grand Canyon has shown that big galleta, bush muhly, and Reverchon threeawn (Aristida purpurea var. nealleyi) can live for more than a century.

A primitive broad-leaved, C3 grass
(Pharus cornutus) found in Coclé, Panama.
The other common growth form in desert grasses is the annual. (Annuals are plants that complete their life cycles — from seed through reproduction and death — in one year or less.) A common trend in grasses and many other desert plants is to evolve toward smaller size and shorter life spans. The best-known adaptations for surviving the heat and drought of fluctuating desert climates are water storage in the succulent stems of cacti, and periodic physiological dormancy in shrubs like creosote bush and brittlebush (Encelia farinosa). However, in avoiding environmental extremes by existing most of the time as seeds in the soil, annual plants have an equally effective adaptation. Often the entire life cycle of desert annuals is compressed into 6 weeks or less; such short-lived plants are aptly called ephemerals.
Desert annuals mostly fall into two groups — winter-spring annuals, related most closely to plants from more temperate zones, and summer annuals, derived from plants of the tropics. Native spring annual grasses include Bigelow bluegrass (Poa bigelovii), little barley (Hordeum pusillum), and sixweeks fescue (Vulpia octoflora). Summer annuals include needle and sixweeks gramas (Bouteloua aristidoides and B. barbata), and panic grasses (Brachiaria arizonica, Panicum hirticaule). Gulf sandbur (Cenchrus palmeri), littleseed muhly (Muhlenbergia microsperma), and sixweeks threeawn (Aristida adscensionis) are opportunistic species that flower in summer or spring. It is interesting that the spring annuals are C3 species derived from temperate relatives, while the summer species, as well as those not restricted to a particular season, are C4s related to grasses of the New World tropics. Sixweeks fescue is widespread in North America from southern Canada to northern Mexico. It reaches its southern limit in tropical deciduous forest in southern Sonora, where winter rainfall from Pacific frontal storms diminishes.
Several desert grasses are at evolutionary stages between perennial and annual. California brome (Bromus carinatus) is a tufted C3 perennial, mostly 15 to 30 inches (40-80 cm) tall, that occurs from Arizona to California and Washington. In Arizona, this perennial was thought to be restricted to elevations of 5500 to 9000 feet (1675 to 2750 m) in the mountains. In 1944, smaller annual plants from the Sonoran Desert near Tucson were described as Bromus arizonicus. Today both plants are considered to be a single species that is perennial in the mountains and annual in the deserts.
Fluffgrass (Erioneuron pulchellum) is a widespread C4 grass in dry rocky slopes up to about 5500 feet (1675 m) from western Texas to Utah and Nevada south into northern Mexico. In summer rainfall climates from the Chihuahuan Desert in Texas west to the northeastern Sonoran Desert near Tucson, it is a tufted perennial no more than 6 inches (15 cm) tall, that mostly flowers in summer or fall, but also sometimes flowers in the spring. On closer examination, individual plants in these populations can be seen to function as either perennials or annuals. Large, 4 to 8 inch (10-20 cm) diameter individuals live in safe sites and last from season to season (perennials). Seedlings colonize open areas in between. These small plants usually do not survive the season, behaving as annuals, but augment seed production and thus chances of recruitment of new individuals into the population. In the Mohave Desert of southern Nevada, fluffgrass is strictly a spring annual.
Sixweeks grama is a widespread summer C4 annual found from Texas and southern Colorado to Arizona and northern Mexico. However, some plants near Tucson are intermediates between sixweeks grama and Rothrock grama (Bouteloua rothrockii), a perennial species endemic to the Sonoran Desert. Both species are part of a closely- related group of gramas with distinctive comb-like flowering spikes, the forms of which range from desert grassland bunch grasses (B. gracilis, B. hirsuta) to desert annuals (B. barbata). Interestingly, the Mayo grama (B. barbata var. sonorae) in southern Sonora is a perennial variety restricted to sandy soils in coastal thornscrub. It proliferates by stolons (above-ground stems that produce new plants, sometimes difficult to differentiate from rhizomes, which are below-ground stems). In this case, the perennial form (as opposed to the desert annual) occurs in more tropical habitats rather than in higher-elevation woodlands and forests. In contrast, dwarf hairy grama (B. quieriegoensis), another southern Sonora thornscrub endemic (it grows nowhere else), is derived from hairy grama (B. hirsuta), a desert grassland species.
Grasses in the Desert
The terms “desert grassland”, “grassland in the desert”, and “grasses in the desert” are sometimes confused.
Desert Grassland
Desert grassland is a distinctive vegetation type in semiarid climates that is widespread at 3500 to 5500 feet (1060 to 1675 m) and is transitional between the short-grass prairie of the Great Plains and desertscrub. The desert grasslands of the Sonoran Desert region are the western edge of the vast grasslands of the middle of the North American continent that extend from the central Canadian provinces south through the Great Plains onto the Mexican Plateau in Chihuahua and Durango, and west across the Continental Divide into New Mexico, southeastern Arizona, and northeastern Sonora. Desert grassland may be dominated by bunch grasses or, especially when disturbed, by various shrubs.
Grasslands in the Desert
Grasslands are found within the Sonoran Desert in very special and interesting ecological situations. In local areas on bajadas and in valley bottoms where clay-rich soils prevent water from draining, grass-dominated communities can develop. At Ventana Ranch between Ajo and Sells on the Tohono O'odham Reservation in Arizona, tobosa (Pleuraphis mutica) covers a periodically flooded valley at 2100 feet (635 m) elevation. This is an isolated, low elevation, western example of the tobosa swales of the Chihuahuan Desert.
Within the Sonoran Desert, from southern Arizona to the broad valleys of central Sonora, the vegetation types change, reflecting milder winter temperatures and modest increases in summer rainfall and relative humidity. The desert changes from the palo verde-saguaro desertscrub of the Arizona Upland (a subdivision of the Sonoran Desert) to the Plains of Sonora (another subdivision), where trees such as desert ironwood (Olneya tesota), palo brea (Cercidium praecox), and tree ocotillo (Fouquieria macdougalii) are common, although columnar cacti are not. At times and in places, grasses are common and important in the vegetation. With the monsoonal rains of July and August, two summer annuals, needle grama and sixweeks grama, flourish in the open spaces between trees and shrubs, especially in the area from Hermosillo to Guaymas. In September and October, their dried stems provide a subtle light yellow fall color to the desert slopes. Although the gramas are visually conspicuous, their slender stems rarely provide enough fine fuel to support a fire, unlike some non-native species (see ?Exotic Species and Ecological Threats? on page 274). In northern Sinaloa, the Mayo Indians collect the grama spikelets from ant mounds and mix them with the clay to make their pots less brittle. Their name, saitilla, applies to needle grama and also to the unrelated Spanish needles (Bidens spp.?an annual composite), both with sharp fruits that readily travel in fur or socks.

Rothrock grama (Bouteloua rothrockii)
On the basis of dense grasses evident in old photographs and the historical presence of the Masked Bobwhite (Colinus virginianus ridgwayi), portions of central Sonora near Benjamin Hill and the Altar Valley of southern Arizona were combined in a new vegetation type called ?Sonoran savanna grassland.? The combination of these diverse areas under the new term is confusing and misleading. ?Savanna? is best restricted to seasonally-flooded tropical grasslands with scattered trees or shrubs. In Africa, ?savanna? has been loosely used to describe any vegetation that has both woody plants and grasses, even if the grasses are subordinate or annuals. In Mexico, tropical savannas mostly occur along the Pacific coast from Sinaloa southward. However, near Benjamin Hill in the Plains of Sonora, long-lived trees and shrubs, rather than short-lived grasses, are generally dominant. The lush grass in old photos was most likely Rothrock grama, a short-lived perennial that may be thick and luxurious in a series of wet years, only to virtually disappear during drought. In contrast, true desert grassland with a rich complement of perennial bunch grasses reaches its western limit in the Buenos Aires National Wildlife Refuge in the Altar Valley of Arizona. Other than the bobwhite and such legumes as velvet mesquite (Prosopis velutina) and white-ball acacia (Acacia angustissima), the Arizona and Sonoran areas in ?Sonoran savanna grassland? share few similarities in either community composition or structure.
The Benjamin Hill area does have local areas dominated by false grama?essentially an endemic type of ?desert grassland.? False grama is a dwarf, tufted perennial grass that spreads prolifically by stolons and forms surprising turfs in open areas. In wetter summers it becomes thick and 6 inches (15 cm) tall. In drought it survives as minute root crowns at the soil surface. The Spanish common name zacate borreguero (?sheep grass?) reflects its ability to withstand heavy grazing by cattle and sheep. False grama is widespread and common in Sonora but has only been found in Arizona in a single population near Raggedtop in the Avra Valley west of Tucson. With selection for greater cold tolerance, it has potential as a low-water use grass in urban landscapes in Arizona. In the hyperarid lowlands of the Lower Colorado River Valley subdivision of the Sonoran Desert in southwestern Arizona, southeastern California, and the Gran Desierto of northwestern Sonora, the vegetation is a simple desertscrub dominated by creosote bush, white bursage (Ambrosia dumosa), and big galleta. In some of the windblown valleys in the Pinacate Region of Sonora, sand piles up on the downwind sides of barren volcanic mountains and big galleta forms pure stands in a starkly beautiful desert landscape.
Grasses in the Desert
In the Sonoran Desert, the grasses are very diverse; over a dozen of them reach the western limits of their ranges in Sonoran desertscrub in the Tucson Mountains. Examples include bull grass (Muhlenbergia emersleyi), cane beardgrass (Bothriochloa barbinodis, formerly called Andropogon barbinodis), curly mesquite grass, gramas (Bouteloua chondrosioides, B. eriopoda, B. gracilis, B. hirsuta), plains bristlegrass (Setaria macrostachya), plains lovegrass (Eragrostis intermedia), and squirrel tail (Elymus elymoides, formerly called Sitanion hystrix). Studies of black grama (B. eriopoda) in the Jornada del Muerto in the northern Chihuahuan Desert in New Mexico suggest that successful reproduction has only occurred during a few periods of unusually wet years in the last century; favorable conditions for reproduction most likely occur even less often on its western range limits.
In addition, a suite of arid-adapted grasses live in the desert communities in Arizona and Sonora, including Arizona cottontop, bush muhly, fluffgrass, tanglehead, and four different threeawns: Aristida californica, A. parishii, A. purpurea, A. ternipes. Grasses are the second most common family (15 percent, 92 taxa) in the Tucson Mountains (610 species total), a desert range usually dominated by palo verde-saguaro desertscrub.
A number of grasses are endemic to the Sonoran Desert. Some of them are widespread from southern Arizona and southeastern California southward into Sonora and Baja California, for example, the threeawns Aristida californica, A. glabrata, and A. parishii, and Rothrock grama. Chloris brandegeei is an annual found in Baja California and on several islands in the Gulf of California. Aristida peninsularis and Bouteloua annua only occur in Baja California. The Gulf sandbur is an annual endemic to sandy habitats in Baja California and Sonora on both sides of the Gulf of California. Sandburs (the genus Cenchrus) have bristles on the fruit that are fused at their bases and modified into the spiny burs that allow the seed to be very effectively transported from place to place by mammals?including humans. Sandburs are closely related to grasses in the genus Cenchrus, which do not have their bristles fused. (Buffelgrass, an exotic species discussed below, is confusing in this regard, since it has weakly-fused bristles; it is placed in either genus by different specialists.) Chromosome evidence indicates that sandburs evolved from Cenchrus.
Desert needlegrass (Stipa speciosa) is one of the few C3 spring-flowering perennial grasses in the Sonoran Desert. It occurs from Colorado to Arizona and southern California. In the Mohave Desert of western Arizona, it flowers in the spring, and is occasionally encountered in desert mountains, where it reaches its southern distributional limit. Fossils in ancient packrat (Neotoma sp.) middens demonstrate that the range of desert needlegrass expanded southward in the winter-rainfall dominated climates of the late Wisconsin glacial period, and especially the early Holocene, 11,000 to 9,000 years ago (see page 63). This grass also occurs in southern South America, as do a number of other species with amphitropical discontinuous distributions (that is, occurring on both sides of the tropics, but not in them?for example, in both North and South America). Other amphitropical species are creosote bush, little barley (Hordeum pusillum), allthorn (Koeberlinia spinosa), and Atamisquea emarginata.
Tanglehead (Heteropogon contortus) is widespread in tropics of the world, a distribution pattern shared by hopbush (Dodonaea viscosa). Tanglehead is an example of a tropical species reaching its northern limit in desert grassland in southwestern United States at about 5500 feet (1675 m) elevation. It is ?wedged out? by cold temperatures at higher elevations and by drought in the desert below. Other tropical species with similar distributions are coral bean (Erythrina flabelliformis), kidneywood or palo dulce (Eysenhardtia polystachya), brown vine snake (Oxybelis aeneus), and green ratsnake (Elaphe triaspis).
Threeawns, genus Aristida, are a taxonomically difficult group containing about 150 species in temperate and subtropical areas in North America and about 40 species in the United States. The fruit usually has 3 awns (stiff hairs, or bristles), thus the common name. The threeawns in the Sonoran Desert present several interesting patterns. Sixweeks threeawn is found from Missouri and Texas west to California and southward to Argentina, and also in warmer parts of the Old World. It is a highly adaptable and opportunistic annual that grows in both spring and summer and can be from 2 inches to 2 feet (50-600 mm) tall, depending on rainfall. Purple threeawn (Aristida purpurea) is a common perennial in Arizona Upland desertscrub in Arizona. Typical purple threeawn (A. p. var. purpurea) with striking, nodding red infloresences commonly occurs on roadsides. (The edges of roads and highways are pseudo- riparian habitats with disturbed soils and concentrated runoff water?easy dispersal corridors for many native and introduced grasses.) If trimmed at the end of the spring and summer growing seasons, purple threeawn is an attractive ornamental in xeriscapes.
In contrast, Reverchon threeawn (A. p. var. nealleyi) only occurs in rocky desertscrub habitats and rarely on roadsides. Superficially it is very similar to A. parishii, which lives in the same upland habitats, mostly differing in details of the fruit. In this threeawn and in many other desert plants, including borages (Amsinckia spp., Cryptantha spp., Plagiobothrys spp.), spiderlings (Boerhavia spp.), and spurges (Euphorbia spp.), there has been far more evolution of the seeds and fruits than of the vegetative plant parts, likely reflecting selection for differences in dispersal methods or microhabitats. In Aristida ternipes, A. t. var. gentilis has 3 awns (bristles) and mostly occurs in desert grassland, whereas spider grass (A. t. var. ternipes) is a single-awned ?threeawn? that is widespread in desert and tropical lowlands. Spider grass is very responsive to rainfall and, like many perennial grasses, can flower in its first year. A seedling encountered in early September in the Tucson Mountains was flowering only 5 weeks after germination in response to a 1.8 inch (46 mm) rainfall. In the desert, spider grass is a low, rounded bunch grass 1 or 2 feet (30-60 cm) tall. In a tropical deciduous forest near Alamos in southern Sonora, with annual rainfall of 20-25 inches (500-630 mm), roughly twice that of the Tucson area, spider grass grows to be an erect, elegant 4 to 5 foot (120-150 cm) tall plant.
Exotic Species and Ecological Threats
There are introduced exotic plants throughout the Sonoran Desert region in Arizona and Sonora. With the notable exception of riparian habitats in Arizona and northern Sonora, introduced species usually account for relatively low percentages of local species present, and are mostly innocuous with few serious impacts on the vegetation. Introduced species are most diverse and abundant in riparian habitats (river bottoms, arroyos, washes) and pseudoriparian habitats (road edges) because they are disturbed, unstable dispersal corridors that harvest water, nutrients, and seeds from large areas. Successful invaders are often short-lived, fast-growing plants, which have high reproductive effort. Longer-lived exotics are usually ?mortality resistant??survivors not easily killed by environmental stresses (floods, fire, drought, freezes, heavy grazing).
Unfortunately a few exotic species have the potential to cause ecological havoc in the Sonoran Desert region and threaten irrevocable landscape changes; most of the worst of these are grasses. Competition with native species is typically intense and can alter the composition of the flora and vegetation. When a new ecological process is introduced into an ecosystem as a result of an introduction of a new species, the impact on the vegetation is more severe?the entire vegetation type can be converted to a different assemblage.
In subtropical desertscrub, tropical thornscrub, and deciduous forest, fire fueled by exotic grasses can be devastating, as most of the community dominants have little or no adaptation to fire.
Some annual grasses native to the Mediterranean areas of Europe are especially troublesome. They are preadapted to the winter rainfall climate and fire regimes of the chaparral vegetation in California. As they have moved eastward into the Mohave Desert, with its winter rainfall, they have directly competed with the native spring flora and introduced fuel and structure conducive to fire.
As they moved further eastward and southward into the biseasonal climatic regimes of the Sonoran Desert, their ecological interactions have been more complex. In desert grassland, competing subshrubs (winter-spring active) and perennial grasses (summer active) have differing water-use strategies. In desertscrub, a similar seasonal competition for soil nutrients occurs between spring and summer annuals. In the spring, introduced annuals compete directly with native spring herbs for water, space, and nutrients. The roots of the introduced annual grasses, including mouse barley (Hordeum murinum), red brome (Bromus rubens), and wild oats (Avena fatua), are active at relatively cool soil temperatures, accelerating their growth compared to native annuals. Often they are so prolific that few nutrients remain for summer ephemerals. However, the alterations of community structure and competition due to fires are much more serious.
Red brome is a weedy Mediterranean annual that was established in California by 1848, and is presently common through much of western United States. It is seasonally abundant and widespread in the Sonoran Desert region. In lower, more arid areas, it is mostly found in disturbed habitats, while at higher elevations throughout the Arizona Upland it also occurs in undisturbed habitats. Observations in a Lower Colorado River Valley creosote bush desertscrub at 1560 feet (475 m) elevation in the Eagletail Mountains west of Phoenix illustrate the variability of these annuals. In the spring of 1992, red brome a foot (30 cm) tall covered the slopes in response to excellent winter-spring rains. The spring of 1996 was very different. In response to a light rain in late February, tiny, approximately 2 inch (50 mm) tall sixweeks threeawn were widespread, but red brome did not germinate, suggesting that native species have some advantages when rainfall is light and soils are relatively warm. Nonetheless, in the last decade or so, early summer wildfires in Sonoran desertscrub, mostly fueled by dried red brome, have increased 10- to 20-fold.
Arabian grass (Schismus arabicus) and Mediterranean grass (S. barbatus) are closely-related winter ephemerals that are geographically segregated in the Old World: Arabian grass from southwest Africa to the western and northern Sahara and the western Mediterranean region, and Mediterranean grass from Kashmir and southern Russia west to Greece. The differences between them are subtle and these plants are difficult to tell apart.
During years of favorable winter rains, these grasses can be abundant across much of the northern part of the Sonoran Desert, forming extensive, dense carpets on relatively flat, sandy terrains. The first stems and leaves often spread out close to the ground, effectively excluding or preventing other ephemerals from sprouting. The earliest record for Arabian grass in North America seems to be from 1933. Mediterranean grass does not seem to be as common in our region as Arabian grass, and apparently does not extend much south of the Arizona border. Interestingly, the earliest North American records for this grass are from 1926 in southern Arizona and from 1935 in Fresno County, California?the reverse of the typical dispersal pattern of Mediterranean weeds, which usually first settle in California and follow the highways eastward. Today this exotic grass is a preferred food for desert tortoises (Gopherus agassizii) in the Mohave Desert, instead of native annuals such as lupines (Lupinus spp.), and deer vetch (Lotus spp.). This may cause physiological problems, because tortoises in the Mohave Desert may need the higher water and protein content of the native annuals to survive the dry summers. In the Tucson Mountains, the introduced red brome joins native curly mesquite grass and fluffgrass as a preferred food of the desert tortoise.
Buffelgrass is a shrubby savannah grass native to the warmer parts of Africa, Madagascar, and India, that is widely introduced and established in hot, semiarid regions of the world for forage and fodder. A strain from the Turkana area of Kenya was officially released for planting by the U.S. Soil Conservation Service in San Antonio, Texas, in 1946. The shrubby habit of buffelgrass allows rapid growth of leaves from the nodes and copious seed production in response to rain. It is very adaptable, likely due to its great genetic variability and largely apomictic reproduction. (Apomixis is a form of asexual reproduction where seeds are produced without fertilization or any sexual union.) The plant burns readily (even when green) and recovers quickly from fires.
Since the 1960s buffelgrass has been extensively introduced for livestock forage in Arizona and Sonora. In Arizona the expansion was initially slow in areas where buffelgrass was not actively seeded into cleared areas. However, in recent years, buffelgrass has begun to spread rapidly along highway shoulders both in lower and higher elevations. It also is invading desertscrub communities on rocky slopes away from roads. In 1992 when a floristic survey of the Tucson Mountains was completed, buffelgrass was only occasionally encountered, and only noticeably invasive on Tucson?s Tumamoc Hill and ?A? Mountain. (On ?A? Mountain, fires accidentally set during the annual Fourth of July fireworks stimulated buffelgrass expansion.) In the past 6 years, dense patches of buffelgrass have become established in several new areas. On Radio Towers Peak, a dirt road provided a dispersal corridor and entry into palo verde-saguaro desertscrub, while on Panther Peak buffelgrass established itself away from roads. In metropolitan Phoenix, buffelgrass is well established on North Mountain and on many other mountains, especially on upper south-facing slopes, where it apparently outcompetes and eliminates native cacti and trees. Such upper slopes appear sandy-colored in winter when buffelgrass is dormant, and are readily distinguishable to the informed eye.
In central Sonora, more than a million hectares (470 thousand acres) of desertscrub and thornscrub have been cleared to plant buffelgrass, often as part of government-subsidized programs to support the ranching industry. Fire cycles in buffelgrass allow the grass to expand rapidly into large areas of uncleared vegetation, very much like cancer spreads in a human body. Along many highways, especially in central Sonora, dense monocultures of buffelgrass have replaced other weedy roadside species, especially brittlebush. Recurrent fires maintain the buffelgrass. Ranchers have been forced to replace wooden posts with metal ones, and the bases of power poles are shrouded with metal or cement. Fires fueled by buffelgrass are now a serious urban problem in Hermosillo, Sonora.
In southern Sonora, buffelgrass has been planted in clearings in tropical deciduous forest but apparently is not able to invade undisturbed shady forests. With management using fire, buffelgrass praderas (prairies) can be maintained indefinitely. However, ranchers are reluctant to use fire lest they lose the last tuft of forage. In many areas, native species including boat-thorn acacia (Acacia cochliacantha), Indian mallow or pintapan (Abutilon abutiloides), and Sonoran bursage (Ambrosia cordata) invade buffelgrass pastures. Without fire, boat-thorn acacia grows tall enough to shade buffelgrass, and succession backwards to tropical deciduous forest may well occur.
This ?grasslandification? of desert-scrub and thornscrub now occurring is the opposite of the well-known ?desertification? of the southwestern United States?the expansion of shrubs into desert grassland beginning in the late-19th century. Ironically, because buffelgrass is such a hardy, drought-resistant grass and excellent forage, great effort has been expended to introduce it and to select hardier varieties. The ecological result of buffelgrass introduction into fire-intolerant subtropical and tropical communities is a permanent (on our time scale) conversion to a savanna, much like those in Africa, with drastically reduced plant cover and overall diversity.
Fountain grass (Cenchrus setaceus) is another robust, often 3 foot (1 m) tall perennial clump grass from Africa. It is a common landscape ornamental in southern Arizona, where it is slowly spreading into natural habitats, especially in desert riparian canyons. It has been cultivated in Tucson since 1940 and was established in the nearby Santa Catalina Mountains by 1946. The larger, purplish cultivar ?Cupreum? is a better choice for landscape plantings, since it does not volunteer, that is, invade areas where it is not planted.
Natal grass (Rhynchelytrum repens) is another African grass that is common on roadsides in many areas in Sonora. This attractive grass with fluffy, rose-colored inflorscences is invasive in desert grassland in Sonora south of Nogales and near Maycoba in the Sierra Madre Occidental, as well as eastward into Chihuahua. In the mountains north of Guaymas on the Gulf of California, it has invaded steep slopes in undisturbed palm canyons. High humidity likely helps it survive in the arid Central Gulf Coast desertscrub. In these conditions, it could potentially fuel fires in vulnerable desertscrub vegetation.
The introduction of the South African Lehmann lovegrass (Eragrostis lehmanniana) has begun a transformation of desert grasslands in southeastern Arizona. It produces 2 to 4 times the annual biomass of native grasses and responds favorably to grazing and fire. It is replacing native grasses in vast areas, dramatically altering community composition without conversion to desertscrub. It has followed major highways well into the Sonoran Desert west of Tucson.
Grasses are one of the largest, most diverse families of plants, found worldwide in virtually all climatic regimes and vegetation types. They are the most important plants economically, providing edible grains for people and forage for livestock. Superbly adapted to live in variable desert environments, they avoid extremes in drought and heat through life history strategies. Well-hydrated plants grow rapidly in response to spring or summer rainfall, dying back to stem nodes (shrubby grasses), root crowns (bunch grasses), or seeds (annuals) during dry periods. The same adaptations help grasses to survive and recover rapidly from fires, grazing, and other severe disturbances.
Some of the many exotic species reaching North America in today?s intercontinental travel and trade are grasses. Successful species from other arid and semiarid lands, especially Africa and the Mediterranean, are rapidly expanding in the Sonoran Desert region, competing with native species. When new ecological processes such as fire are introduced, the impacts are devastating. Fire-intolerant desertscrub and thornscrub can be permanently converted to Africanized ?savannas? or weedy Mediterranean annual ?grasslands.? Increases in fires and in the total area of disturbed communities in the last few decades in Arizona and Sonora should be a call to arms to battle these invasive Old World grasses.
Additional Readings
Gould, Frank W. Grasses of Southwestern United States. Tucson: University of Arizona Press, 1977.
McClaran, Mitchel, and Thomas R. Van Devender, eds. The Desert Grassland. Tucson: University of Arizona Press, 1995.
Webb, Robert H. Grand Canyon: A Century of Change. Rephotography of the 1889-1890 Stanton Expedition. Tucson: University of Arizona Press, 1996. (Included here is a table of plants recorded to live more than a hundred years, including grasses.)

