






The Deep History of the Sonoran Desert
The Sonoran Desert is considered to be the most �tropical� of the North American deserts. Its climate is virtually frost-free, and summer rainfall comes from the tropical oceans. The Sonoran Desert�s structurally diverse vegetation, which includes columnar cacti and leguminous trees, certainly differs from those of the shrub-dominated Great Basin, Mohave and Chihuahuan deserts. It has both geographic and biologic connections with more tropical communities. In a single day�s travel, naturalists can begin in the oak woodlands and desert grasslands in southern Arizona along Interstate 19, and travel through the various desertscrub, thornscrub, and tropical deciduous forest habitats along M�xico 15 in Sonora, experiencing the remarkable transition from temperate zone communities to the New World tropics. This vegetational gradient gives us a sense of the Sonoran Desert�s connection with the tropics, but does not really explain it. The explanation lies in the tropical roots of the Sonoran Desert, deep in its evolutionary history.
A Walk through Time
The Sonoran Desert that we see today, with its characteristic assemblages of plants and animals is quite recent, at least in terms of geologic time. In
fact, it and the other North American deserts are among the youngest
biotic communities on the continent. Although some Sonoran species evolved in ancestral seasonally-dry tropical communities, the development of the unique regional climates and the evolution of characteristic desert-adapted plants and animals are thought to have combined to form the Sonoran Desert by about 8 million years ago (mya) in the late Miocene. Similar conditions developed many times subsequently
as global climates changed, with the Sonoran Desert continually expanding, contracting, and redefining itself. The most recent expansion of the Sonoran Desert into its modern area in Arizona and California occurred only 9000 years ago, with the modern communities of plants and animals developing 4500 years later. This chapter is a walk through time examining the conditions that led to the development of the Sonoran Desert and exploring what shaped its dynamic history. ![]()
The Paleocene
(66.4 to 57.8 mya)
In the Paleocene epoch, soon after the extinction of the dinosaurs (65 mya), most of North America was covered with temperate evergreen and tropical rainforests. There was little regional variation. The warm climates promoted humid forests with strong Asian affinities; primitive ferns (Anemia spp.), cycads (Di�n, Zamia spp.), and palms grew as far north as Alaska. The flowering plants (angiosperms), whose spectacular evolutionary radiation began in the Late Cretaceous, became increasingly important in the forests, displacing archaic cycads, conifers, and tree ferns. The
earliest indisputably recognizable fossil grasses were found in sediments dating to about 58 mya. These were broad-leaved forest grasses ancestral to the modern bamboos. ![]()
The Eocene
(57.8 to 36.6 mya)
In the Eocene, deciduous trees became increasingly common, providing the first evidence of a dry season. The landscape now included tropical deciduous forests, in which trees dropped their leaves in response to drought. During these periods, sunlight passed through the leafless canopy and heated and dried the surface of the ground. Many new species of plants and animals evolved, adapting to these new heat and moisture regimes. The origins of cacti and other succulents likely occurred during the Eocene in dry tropical forests. Fossils of an alligator (Allognathosuchus), a softshell turtle (Trionyx), a primitive tortoise (Geochelone), a primitive monitor lizard (Varanidae), a ground boa (Boidae), and many small mammals from Ellesmere Island in northeastern Canada, then at 78� latitude, indicate that the world was very warm and that plants and
animals freely traversed a land bridge between North America and Europe. The results were dramatic shifts in
the biota as more advanced forms
displaced archaic ones. ![]()
The Oligocene
(36.6 to 23.7 mya)
By the Oligocene, grasses, including the important taxonomic groups in arid lands, had achieved relatively
modern diversity. Unfortunately,
most of this dramatic radiation in the grasses, one of the most important plant families, was not captured in the fossil record. Gopherus, the genus of the modern desert and gopher tortoises
(G. agassizii and G. polyphemus), appeared in the Oligocene. Modern genera of lizards in the Oligocene fauna were skinks (Eumeces) and beaded lizard or Gila monster (Heloderma). The snake fauna was dominated by small ground boas related to the living rubber boa (Charina bottae) and desert rosy boa (Lichanura trivirgata) of western North America and the sand boas (Eryx spp.) of Africa. The lizards common today in the Sonoran Desert�Iguanidae and Teidae, (that is, iguanas and their relatives, and whiptails)�and the common snakes of the Sonoran Desert today�Colubridae and Viperidae (colubrid snakes, for example, bull snakes, king-snakes; and pitvipers) were uncommon or absent at these early dates. ![]()
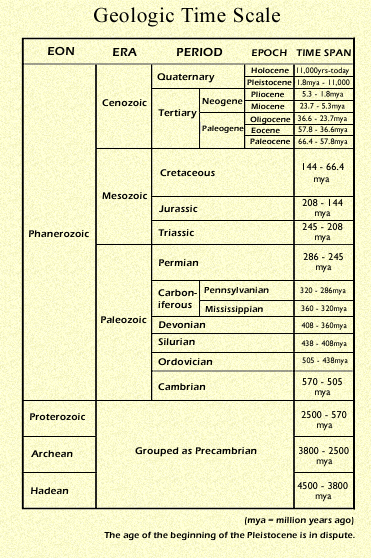
The Miocene
(23.7 to 5.3 mya)
A series of enormous volcanic eruptions from the middle Oligocene to the middle Miocene (about 30 to 15 mya) changed the climates and established
the modern biogeographic provinces
of North America. (See the chapter �The Geologic Origin of the Sonoran Desert� for a more complete discussion of these processes and the terminology used to describe them.) The Rocky Mountains were uplifted to new heights by the accumulation of over a kilometer and a half of volcanic rock. A kilometer (.6 mile) thick layer of rhyolitic ash fell in the Sierra Madre Occidental in northwestern Mexico�on top of a kilometer of early Tertiary (the geologic period that includes the Paleocene, Eocene, Oligocene, Miocene, and Pliocene epochs) andesites. As regional uplift pushed them even higher, the mountains interrupted the upper flow of the atmosphere for the first time. Tropical moisture from both the Pacific Ocean and the Gulf of Mexico was blocked from the mid-continent, drying out the modern Great Plains and the Mexican Plateau. Harsher climates
segregated drought- and cold-tolerant species into new environmentally-
limited biomes, including tundra, conifer forests and grasslands, and restricted them along elevational and
latitudinal environmental gradients.
The Miocene also was a time when major evolutionary radiations began
in many of today�s successful groups including composites (plants in the
sunflower family), grasses, toads,
iguanid and teid lizards, colubrid
snakes and pitvipers. ![]()
The �Miocene Revolution�
After the rise of the Sierra Madres and the resulting changes in climate, tropical forests were found only in the lowlands along the coasts of Mexico and Central America. There, newly evolved species joined archaic ones in the biome known as tropical deciduous forest. And, along the lower, drier edges of tropical deciduous forest evolved a new biome�thornscrub. Thornscrub looks very much like a transitional state between tropical deciduous forest and Sonoran Desert: its vegetation is shorter and sparser than tropical forest and it does not require much moisture. During the Miocene, thornscrub may well have been the regional vegetation in drier areas to the north that are now Sonoran Desert. Thornscrub, in fact, may be the ancestral biome of many Sonoran Desert plants and animals.
The Sonoran Desert itself came about during a drying trend in the middle Miocene (15 to 8 mya). Much of the desert�s vegetation, however, predates the Sonoran Desert itself, having evolved in climates that also required adaptations to aridity. For example, guayac�n (Guaiacum coulteri), organpipe cactus (Stenocereus thurberi), palo brea (Cercidium praecox), senita (Lophocereus schottii), and tree ocotillo (Fouquieria macdougalii) likely evolved in thornscrub. Other plants such as desert ironwood (Olneya tesota), foothills palo verde (Cercidium microphyllum), and saguaro (Carnegiea gigantea) evolved along with the Sonoran Desert.
Another important chapter in
the history of the Sonoran Desert
concerns the formation of the Baja California peninsula. Before 12 million years ago, much of the land which is now Baja California was part of the Mexican mainland. Activity along the San Andreas fault caused the Gulf of California to open, and several large chunks broke away from the mainland and drifted in splendid isolation northwestward. The timing of the formation of Baja California is controversial, with some estimates as recent as 5 to 6 million years ago. These islands were populated with tropical plants and animals which soon evolved into regional endemics, including the boojum tree or cirio (Fouquieria columnaris). Eventually the islands combined and joined California to form the Baja peninsula. Today, the modern islands on both sides of the peninsula in the Pacific Ocean and the Gulf of California, many of which are geologically young, are the most active evolutionary arenas in the Sonoran Desert Region, and have many endemic species. ![]()
The Pliocene
(5.3 to 1.8 mya)
During the latest Miocene and early Pliocene, geological forces again altered landscapes and climate regimes, causing a reversal to more tropical climates.
Sea level rose enough that the Gulf
of California expanded into the Los Angeles area of southern California.
A fossil skull of an iguana (Pumilia novaceki), a primitive relative of the
tropical green iguana (Iguana iguana)
that today occurs no farther north
than southern Sinaloa, was found in sediments 2.5 to 4.3 million years old in southern California. With tropical circulation patterns enhanced by warmer oceans, tropical forests of western Mexico likely expanded, reaching farther north than they do today in Sonora; likewise, the Sonoran Desert in Arizona and California extended further, perhaps as far as southern Nevada. ![]()
The Pleistocene
(1.8 mya to today)
The warmth of the Pliocene ended abruptly at the beginning of the Pleistocene about 1.8 million years ago, as the Earth entered a new climatic era that far surpassed the middle Miocene in cool, continental conditions. Traditionally, four ice ages or glacial periods were recognized, based on terrestrial sedimentary deposits in North America, and these were widely correlated with conditions in Europe and South America. However, recent studies of sediment cores from the ocean floors record fifteen to twenty glacial periods in the Pleistocene. Ice ages were about ten times longer than interglacials (the warm periods between ice ages) which lasted 10,000 to 20,000 years. Officially, the end of the Pleistocene was defined as the beginning of the Holocene 10,000 years ago, based on changes in sediments in European lakes. Today we understand that the Holocene is the present interglacial period and that the cyclic environmental fluctuations of the Pleistocene likely have not ended.
In the last glacial period (the Wisconsin), the massive Laurentide continental glacier covered most of Canada and extended as far south as New York and Ohio. As much as two miles (3 km) of ice covered the Great Lakes and New York City. Boreal forest with spruce and jack pine moved southward displacing the mixed deciduous forests of the eastern United States. Mountain glaciers covered the tops of the Rocky Mountains and the Sierra Nevada in the western United States and the Sierra Madre del Sur in south-central Mexico. Now-dry playa lakes in the Great Basin were full. Enough water was tied up in ice on land to lower sea level about 425 feet (130 m).
During the last half of this glacial period (from 45,000 to 11,000 years ago), plant remains in ancient packrat (Neotoma spp.) middens document the expansion of woodland trees and shrubs into areas that had been desert. Woodlands with singleleaf pinyon (Pinus monophylla), junipers (Juniperus spp.), shrub live oak (Quercus turbinella), and joshua tree (Yucca brevifolia) were widespread in the present Arizona Upland subdivision of the Sonoran Desert. Ice age climates with greater winter rainfall from the Pacific Ocean and reduced summer monsoonal rainfall from the tropical oceans favored woody cool-season shrubs related to plants living farther north, rather than to the summer-rainfall trees, shrubs and cacti of tropical forests and subtropical deserts. The isolated chaparral communities in central Arizona, mostly in a northwest-southeast band below the Mogollon Rim, are relicts of ice-age chaparral connections with California. Many species are shared between California and Arizona chaparral, including shrub live oak. The Arizona black rattlesnake (Crotalus viridis cerberus) in Arizona chaparral is essentially a dark form of the southern Pacific rattlesnake (C. v. helleri); it is more distantly related to the other four subspecies of western rattlesnakes in Arizona.
Warm desert communities dominated by creosote bush (Larrea tridentata) were restricted to below 1100 feet (300 m) elevation in the Lower Colorado River Valley in the Sonoran Desert and in the southern Chihuahuan Desert. Although brittlebush and saguaro returned to Arizona soon after the beginning of the present interglacial (the Holocene) about 11,000 years ago, the Sonoran Desert did not re-form until about 9000 years ago, as the last displaced woodland plants retreated upslope. Relatively modern community composition was not achieved until about 4500 years ago when foothills palo verde, desert ironwood and organpipe cactus arrived from their retreats to the south and to low elevations. However, the modern assemblages that we recognize as the Sonoran Desert communities must have recurred many times during the Pleistocene interglacials, only to retreat to warmer
climates as ice age climates returned. Modern desert communities have been present for only about five percent of the 2.4 million years of the Pleistocene, while ice age woodlands in the desert lowlands persisted for about ninety percent of this period. ![]()
Tropical Interglacials
Surprisingly, the vertebrate fossil record suggests that some interglacial climates were more tropical during the Holocene. El Golfo de Santa Clara is near the mouth of the Colorado River in northwestern Sonora. Early Pleistocene (1.8 mya) fossils reflect a climate that was frost free, with much greater rainfall in the warm season, and with higher humidity than today. Greater summer rainfall would suggest that tropical oceans were warmer than they are today, in contrast to most of the Pleistocene when ocean waters were colder. The fauna included such mammals as antelope, bear, camels (dromedaries and llamas), cats, horses, proboscidians, and a tapir (Tapirus). The giant anteater, capybara (Neochoerus), and ground sloths in the fauna were members of ten families of mammals that immigrated into North America in the late Pliocene or early Pleistocene after the opening of the Panamanian land bridge during the Great American Interchange. In contrast, the imperial mammoth (Mammuthus imperator), a hyena (Chasmoporthetes johnstoni), and jaguar (Felis onca), were Eurasian immigrants. The nearest populations of giant anteater are 1800 miles (3000 km) to the southeast in the humid, tropical lowlands of Central America! As for many large mammals, the modern distribution may not accurately reflect their physiological range limits because of human predation in the last 11,000 years. Other fossils in the fauna include the Sonoran Desert toad (Bufo alvarius), slider �turtle�(Trachemys scripta), boa constrictor (Constrictor constrictor), and the large extinct California beaver (Castor cf. C. californicus). The Sonoran Desert toad is a regional endemic, while the slider and boa constrictor occur today in Sonora in wetter, more tropical areas to the southeast. Although the El Golfo area is today part of the hyperarid Gran Desierto, the delta of the Colorado River was historically a very wet area that supported extensive cottonwood (Populus fremontii) gallery forests with abundant beaver. There is even a December 1827 account of a large spotted cat (likely a jaguar) that entered James Ohio Pattie�s camp on the Colorado River south of Yuma to feed on drying beaver skins.
Rancho La Brisca is in a riparian stream canyon north of Cucurp�, fifty-four miles (90 km) south of the Arizona boundary in Sonora. A pocket of 150,000-year-old ci�nega sediments yielded abundant Sonoran mudturtle (Kinosternon sonoriense), fish, and other small vertebrate fossils associated with bison (Bison). The presence of the sabinal frog (Leptodactylus melanonotus) 144 miles (240 km) north of the northernmost extant population on the R�o Yaqui indicates that the climate of a late Pleistocene interglacial was also more tropical than at the site today. ![]()
Sonoran Desert Mammoths?
A few years ago, a Mayo Indian found a very large bone in the bank of the arroyo behind his house in Teachive, a village in coastal thornscrub in southern Sonora. For him, as it has been for others who have discovered fossil mammoth bones throughout North America and Europe for centuries, it was puzzling and perhaps frightening. What animal could be so much larger than a deer or a cow? Why has no one seen these monsters? Many a legend was born to explain them and their disappearance.
Today we know that about 11,000 years ago, nearly two-thirds of the large mammals of North America went extinct. Common, widespread grazers, including horses and mammoths, disappeared at the very time that spruce and pine retreated and grasslands expanded from Arizona to Canada. Paul S. Martin of the University of Arizona forcefully presented the case that big game hunters caused widespread extinctions within a few hundred years after their entry into North America from Siberia via the Bering Strait. The theory of �overkill� of �naive� large mammals is controversial, and some suggest changes in climate may have caused the extinctions. However, the paleobotanic record gives no evidence of climatic changes severe enough to have resulted in the extinction of so many large animals over such a broad, diverse area. The well-preserved plant remains in packrat middens provide additional insights.
A species could respond to a major
climatic change by (1) adapting
genetically (speciation), (2) becoming extinct, or (3) adjusting its geographical distribution. At the beginning of the Holocene, the last glacial/interglacialclimatic shift, there are essentially no records of speciation or extinction in plants or small animals. Most simply shifted their geographic and elevational ranges. Moreover, woodland plants
survived in desert lowlands for several thousand years after the megafaunal extinctions and before the expansion of the Sonoran Desert. The biotic communities of North America have had fewer large herbivores in the last 11,000 years than at any time in the last 20 million years! The impacts of these herbivores on tropical deciduous forest, thornscrub, and the Sonoran Desert flora were undoubtedly profound�but we will never fully understand the ecological roles of these missing animals. ![]()
The Arctic Connection
Tropical communities with their great species diversity, outrageous morphological adaptations, and mixtures of archaic and advanced species have been important evolutionary arenas. However, the evolutionary mechanisms are not so clear, considering that the climatic fluctuations that isolate populations and stimulate the evolution of new species are more intense at high latitudes. Recent paleomagnetic dating of fossil-bearing sediments in northern Canada indicate that some plants appeared there as much as 18 million years earlier, and some mammals two to four million years earlier than at lower latitudes. If the relative ages are not the result of a fragmentary fossil record, they may indicate that important biotic innovations evolved in the mild �tropical� Arctic climate with its months-long polar day-night cycle, and then the species moved southward into the tropics.
Fossil records of tropical plants and animals from Arctic latitudes with six-month-long nights not only reflect very warm global climates but raise questions about how these organisms survived the dark. Today reptiles
spend the cold winters in hibernation and hot, dry periods in estivation. Deciduous plants shed their leaves for long periods, either triggered by the onset of cold temperatures in temperate latitudes or the beginning of the dry season in tropical latitudes. The Arctic fossils suggest that deciduousness in plants and hibernation and
estivation in reptiles could have arisen as responses to the polar night, and later shifted to these other stimuli. ![]()
Whose Child Is This?
Although the perception of the Sonoran Desert as �tropical� is based partly on the presence of columnar cacti, there are climatic, physical, and deep historical connections as well. As discussed above, the region likely supported tropical deciduous forest from the Eocene to the early Miocene and then thornscrub later in the Miocene. The desert biota of this region are rich in endemics, most of which evolved in more tropical communities prior to the Sonoran Desert itself. Many Sonoran species reach their southern limits in thornscrub�the structural, biotic, and historical link to tropical deciduous forest�not in the tropical deciduous forests of southern Sonora. With the exception of organpipe cacti and a few others, the paucity of desert species shared with the tropical deciduous forests challenges the popular idea that �tropical deciduous forest is the mother of the Sonoran Desert.�
Thornscrub could more accurately be called the �mother� of the Sonoran Desert and tropical deciduous forest its �grandmother� or �great aunt�! ![]()
Additional Readings
Bahre, Conrad J. A Legacy of Change. Historic Human Impact on Vegetation in the Arizona Borderlands. Tucson: University of Arizona Press, 1991.
Betancourt, Julio L., Thomas R. Van Devender, and Paul S. Martin. Packrat Middens. The Last 40,000 Years of Biotic Change. Tucson: University of Arizona Press, 1990.
Davis, Goode P., Jr. Man and Wildlife in Arizona: The American Exploration Period 1824-1865. Phoenix: Arizona Game and Fish Department, 1982.
Grayson, Donald K. The Desert�s Past. A Natural Prehistory of the Great Basin. Washington, D.C.: Smithsonian Institution Press, 1993.
Hastings, James R. and Raymond M. Turner. The Changing Mile. Tucson: University of Arizona Press, 1965.
Imbrie, John and Kathryn P. Imbrie. Ice Ages. Solving the Mystery. Cambridge: Harvard University Press, 1986.
Kurt�n, Bjorn and Elaine Anderson. Pleistocene Mammals of North America. New York: Columbia University Press, 1980.
Martin, Paul S. and Richard G. Klein, eds., Quaternary Extinctions: A Prehistoric Revolution.
Tucson: University of Arizona Press, 1989. ![]()

