Migratory Pollinators Program
 Carnegiea gigantea |
 Carnegiea gigantea |
 Stenocereus thurberi |
 Agave parryi |
 Pachycereus pringlei |
 Pachycereus pringlei |
 Agave cf. wocomahi |
 Agave sp. flowers |
LESSER LONG-NOSED BATS: RESULTS
Roosts and
Surveys
Bat roost data was compiled for roosts throughout the region.
Results for a total of 46 sites can be seen in Table 1. New
maternity roost sites have not been discovered in recent years. During this
study, Karen Krebbs discovered a new maternity site in Cucurpe, Sonora.
The discovery of the site was especially rewarding because it was the result of
a lead from a telesecundaria (satellite middle school) teacher after a
slide presentation on bats to her class.
Carbon
Stable Isotope Analysis
It is likely that nectar and pollen provide
most of the fuel used by lesser long-nosed bats during their spring and fall migrations.
Thus, knowing which floral resources these bats visit, and their flowering phenologies
and geographic distributions, help to define their migratory corridors.
The diet of the lesser long-nosed bat at many locations throughout its range was
examined using carbon stable isotope techniques
(Fleming et al. 1993, Ceballos et al. 1997). This method allows estimation
of the relative contribution of crassulacean acid
metabolism (CAM) plants and Calvin
(C3) plants to the diet of this bat as these two groups of plants differ significantly
in the ratio of 13C to 12C (d13C
) in their tissues (Fleming 1995).
Results of the analyses indicate that the relative importance of CAM plants in the diet of the lesser long-nosed bat varies geographically and temporally in Mexico. Bats from Baja California Sur, for example, are enriched in 13C year round, which indicates that these bats are CAM specialists. In contrast, bats living in Jalisco have low d13C values year round and are feeding primarily on non-succulent (C3) plants. Bats that travel north in the spring change in composition from a mixture of CAM and C3 plants in January and February to CAM plants by April. Bats living in the Sonoran Desert during the summer are feeding wholly on CAM plants. During the fall migration, bats shift back from primarily a CAM plant diet to a mixture of CAM and C3 plants by November and December. Pollen analyses also showed that bats are feeding on columnar Cactaceae almost exclusively (more than 99% of the total pollen sum in most samples were columnar Cactaceae) when they are flowering.
These data indicate that lesser long-nosed bats use CAM plants to fuel their spring and fall migrations.
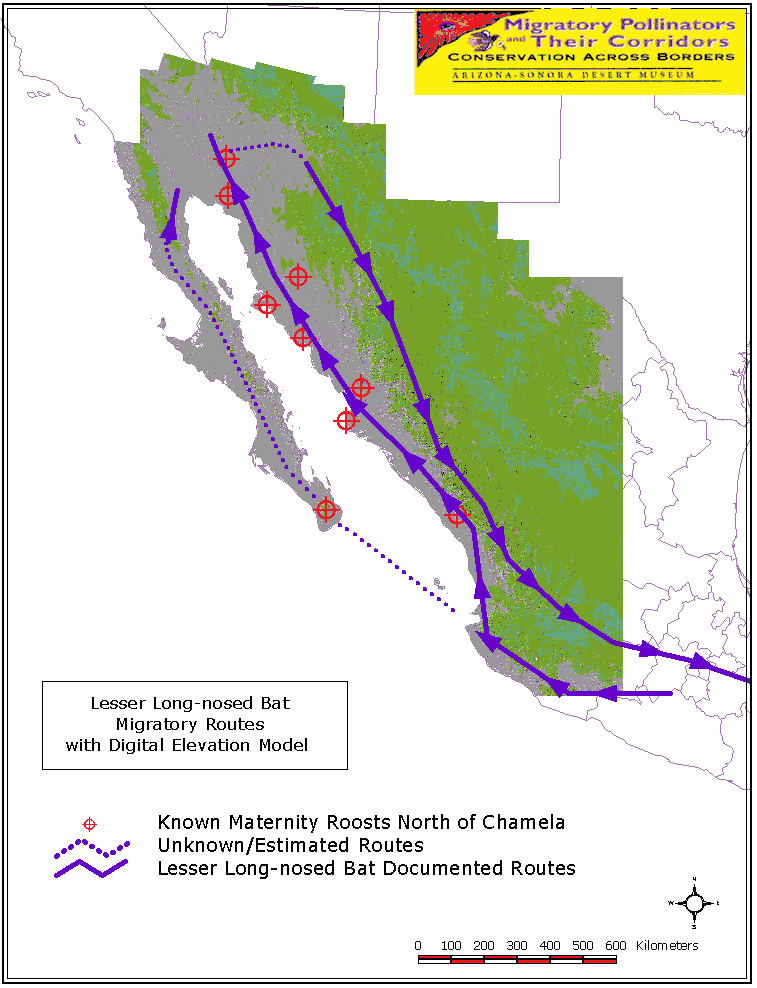
In the spring, these plants include several species of columnar cacti that flower in a northward progression along the coastal lowlands of western Mexico. In the fall, these plants include several species of paniculate agaves that flower in a southward progression in the foothills and western flanks of the Sierra Madre Occidental of Mexico. The flowering phenologies and geographic distributions of these columnar cacti and paniculate agaves suggest that lesser long-nosed bats may migrate into and out of the Sonoran Desert along two different nectar corridors: a coastal cactus corridor in the spring (Map 7) and an inland agave corridor in the fall.
Field observations made throughout the corridors (Map 7) confirm the carbon stable isotope analysis. The columnar cacti used by lesser long-nosed bats during spring migration (and during the brooding period) are saguaro (Carnegiea gigantea), organpipe cactus (Stenocereus thurberi), cardón (Pachycereus pringlei), and etcho (Pachycereus pecten-aboriginum). The agave species used during the fall (southbound) migration is Agave palmeri.
Mitochondrial
DNA (mtDNA) Analysis
The traditional way in which bat biologists have studied
migration is to band thousands of individuals at different localities with the
hope that some of those individuals will later be found or recaptured someplace
else. This is a labor intensive, inefficient method because rates of band
returns are extremely low (< 1%), at least for bats that migrate between Mexico
and the United States (e.g., Cockrum 1969). In the case of the long distance
migratory lesser long-nosed bats, large-scale banding programs are out of the
question. An alternative method for determining seasonal movement patterns
is mtDNA analysis.
MtDNA is especially appropriate for this kind of analysis because it evolves (i.e. undergoes random nucleotide substitutions) more rapidly than DNA (deoxyribonucleic acid), hence it provides more variation for analysis, and it is inherited maternally thus undergoes no recombination. This means that, aside from mutations, it is passed unchanged from generation to generation matrilineally. For our study, tiny samples of wing membrane were collected from individuals at 18 locations throughout Mexico and southern Arizona. When the mtDNA haplotype of each individual was identified, roost sites containing the same haplotypes could be matched up and identified. These genetic data were subjected to a variety of statistical analyses to look for nonrandom patterns of haplotype distributions.
 |
| Map 7. Click for full size map with key |
MtDNA studies revealed two important patterns (Fleming in press). First, haplotypes found in the mating cave near Chamela, Jalisco, also occurred in maternity caves in northern Sonora and southern Arizona as well as in southern Baja California. These data provide direct evidence of seasonal movements by bats of up to 1,200 km between Jalisco and Arizona. Second, the mtDNA data suggest that lesser long-nosed bats probably use two migration paths to reach northern Sonora and southern Arizona each year (Map 7). The first path corresponds to the early spring coastal lowland route suggested by the carbon isotope data. The second path is located farther inland (along the foothills and western flank of the Sierra Madre Occidental) and brings bats from central Mexico into southeastern Arizona in the spring and summer. Some of the bats using this inland route are adult males whereas virtually all of the bats using the coastal route are females. Why these males migrate north is currently unknown.
{kind=link}
Fleming (unpublished data from this project) reports that migrating females are more likely to move within a corridor than between corridors supporting the coastal and montane corridor hypothesis. According to Fleming, female bats do not appear to be moving from coastal maternity sites into montane post-maternity sites. Further research is necessary to determine other potential locations of coastal post-maternity sites. The cladistic analysis of migration suggests that females are divided into at least three groups: spring-birth coastal, spring-birth montane, and winter-birth groups. Analysis also indicated that movement is restricted between winter-birth deme and spring-birth deme for females, but not for males; however, no significant genetic population subdivision was detected between reproductive demes.
Results and conclusions continued on Next Page
Top of Page | Previous Page | Next Page